Bot. Bull. Acad. Sin. (2004) 45: 127-131
Qaddoury and Amssa Effect of IBA on the biochemical changes in relation to root formation in date Palm offshoots
Effect of exogenous indole butyric acid on root formation and peroxidase and indole-3-acetic acid oxidase activities and phenolic contents in date Palm offshoots
Ahmed Qaddoury1,* and Mohamed Amssa2
1Laboratoire de Biotechnologie et Physiologie Végétales, Université Cadi Ayyad, Faculté des Sciences et Techniques - Guéliz, B.P. 549, Marrakech, 40 000 Morocco
2Laboratoire de Biotechnologie et Physiologie Végétales, Université Mly Ismail, Faculté des Sciences, Meknes, Morocco
(Received July 15, 2003; Accepted November 18, 2003)
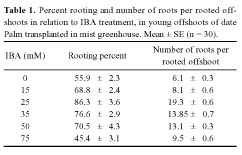
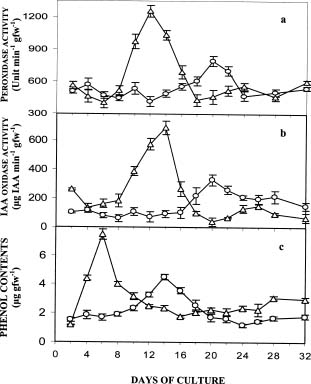
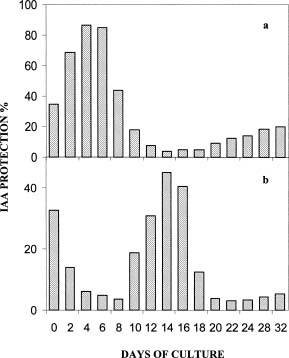
Abstract. Exogenous indole butyric acid had a significant positive effect on the rooting response of date Palm (Phoenix dactylifera L.) offshoots. IBA-treated offshoots rooted earlier and with a much higher frequency while untreated offshoots rooted poorly, later, and tended to produce fewer roots. The effect of exogenous application of auxin was also reflected in the metabolic changes in offshoots during the rooting process. Pox activity showed a minimum on the sixth day and a maximum on the twelfth day after the root-inducing treatment. Changes of IAA oxidase activity showed a parallel time curve while phenolic contents and auxin protector levels showed an inverse trend. These parameters changed slightly one week later in the untreated offshoots.
Keywords: IAA oxidase; IBA; Offshoots; Phoenix dactylifera; Phenolic compounds; Peroxidase; Rooting.
Abbreviations: HRP, horseradish peroxidase; IBA, indole butyric acid.
Introduction
The rooting property of auxins and their importance in plant propagation have been widely recognized for long time (Thiman and Went, 1934). Hence, the necessity of exogenous auxin application to induce root formation in cuttings has been reported in many species e.g. sequoia (Berthon et al., 1990), loblolly pine (Greenwood and Weir, 1994), juniper (Edson et al., 1996), almond (Caboni et al., 1997) and Eucalyptus (Fett-Neto et al., 2001). Currently, indole butyric acid (IBA) is the most widely used auxin to stimulate the rooting process in cuttings because of: 1) its high ability to promote root initiation (Weisman et al., 1988) and 2) its weak toxicity and great stability in comparison to naphthalene acetic acid and indole-3-acetic acid (Blazich, 1988; Hartmann et al., 1990). Though auxins are widely used in the induction of rooting, little is known about their specific action or their interactions with other endogenous compounds (Gaspar et al., 1997). Changes in enzyme activities which regulate different biochemical pathways e.g. protein, carbohydrates, nitrogen, and phenolics, during the rooting process in cutting have been investigated (Mato et al., 1988; Das et al., 1997; Druege et al., 2000). Several investigations have reported the involve
ment of peroxidase, IAA oxidase, and phenolics in the process of root formation in cuttings (Hahlbrock and Grisebach, 1979; Mosella et al., 1980; Vaughn and Duke, 1984; Mayer, 1987; Gonzalez et al., 1991; Caboni et al., 1997; Gaspar et al., 1997).
Date Palm multiplication by transplanting offshoots still remains the best and most common method of reconstituting a destroyed palm grove. Its accomplishment was however limited to offshoots that weigh 7 to 15 kg (Perreau-Leroy, 1958; Toutain, 1972; Saaïdi et al., 1979). In the last few decades, the rooting ability of the date palm offshoot has substantially improved using the misting system (Saaïdi, 1979; BenAbdelah, 1990; Qaddoury and Amssa, 2002). The present investigation was carried out to provide a more detailed insight into the relationship between the auxin (IBA) treatment and changes of peroxidase and IAA oxidase activities as well as endogenous phenolic contents and auxin protector levels in relation to rooting response, in the offshoot of the high quality Moroccan cultivar of date palm.
Materials and Methods
Plant Material
Young offshoots, less than 2 kg, were collected from the high quality Moroccan cultivar of date palm, Mejhoul (MJH), grown in the experimental station of the ORMVAT
*Corresponding author. Tel: (212) 61 25 96 96; Fax: (212) 44 43 31 70; E-mail: qaddoury@fstg-marrakech.ac.ma