Bot. Bull. Acad. Sin. (2004) 45: 133-141
Wahid Toxic and osmotic effects of NaCl on sugarcane
Analysis of toxic and osmotic effects of sodium chloride on leaf growth and economic yield of sugarcane
Abdul Wahid*
Department of Botany, University of Agriculture, Faisalabad-38040, Pakistan
(Received October 7, 2003; Accepted December 2, 2003)
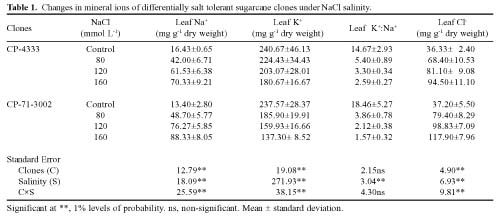
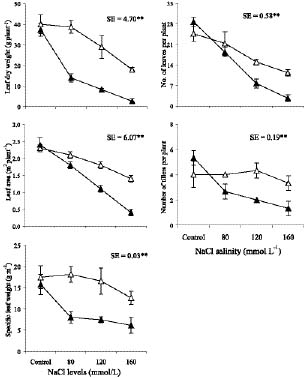
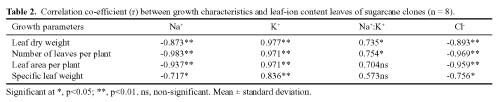
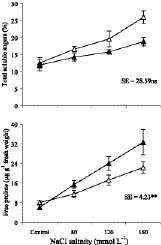
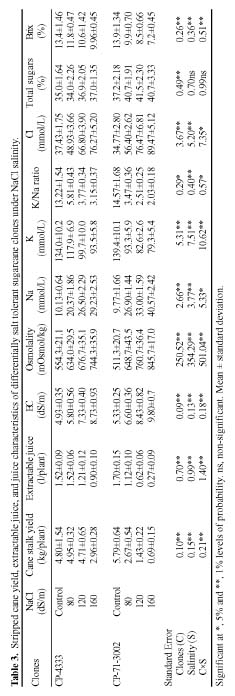
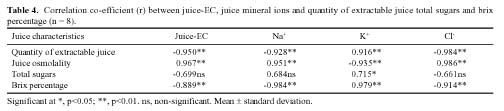
Abstract. Sugarcane (Saccharum officinarum L.) an important sugar crop, shows high sensitivity to salinity at various growth stages. This study was conducted to determine the comparative response of a salt-tolerant (CP-4333) and a salt-sensitive (CP-71-3002) sugarcane clone to the toxic or osmotic effects of sodium chloride at two growth stages. Clones indicated significant differences in terms of reductions in dry weight and area of leaves under salinity at the grand growth stage. Leaf dry weight was more affected than leaf area, resulting in reduced specific leaf weight. The tolerant clone produced supplementary tillers in greater number that appeared to offload the ion excess. Clones indicated significant difference with regard to increase in leaf Na+ and Cl- and decrease in K+, but no difference in K+:Na+ ratio. Na+ and Cl- were negatively correlated while K+ was positively correlated with leaf growth parameters, indicating an adverse effect of Na+ and Cl- and importance of K+ to salt tolerance. The tolerant clone displayed higher water content, water, and turgor potentials of leaf than sensitive clone, but the osmotic potential did not significantly change. Soluble sugars of tolerant clone indicated a ~2 fold increase over control, indicating their osmo-protective role. Free proline accumulation was more specific to the sensitive clone and was correlated with Na+ and Cl- showing its synthesis due to ion-toxicity. At maturity, increased salinity reduced the millable cane yield, extractable juice and juice-brix percentage, but increased juice osmolality. All these parameters were negatively correlated with EC, Na+, and Cl- and displayed their specific effect on the sugar levels in the internodes. In summary, ion-toxicity was the main determinant of salt tolerance at the grand growth stage while the osmotic component of NaCl mainly appeared to affect the transport of sucrose to stalks, followed by stimulated sucrolytic activity in the internodes, resulting in reduced final cane yield.
Key words: Brix; Internodes; Invertase; Ion-toxicity; K+; Specific leaf weight; Sugars; Tillering; Turgor.
Introduction
A soil is considered saline or salt-affected when the electrical conductivity of extract from water-saturated, root-associated soil exceeds 4 dS m-1. Salt affected land on earth comprises 19% out of 2.8 billion hectares of arable land (Szabolics, 1989). The main reason for this in irrigated areas is the build-up of excess ions in the upper soil profiles while in non-irrigated areas the culprit is a low precipitation/evaporation ratio. The build-up of toxic ions in the rhizosphere initially causes injury to plant roots, and then their gradual accumulation in the aerial parts causes heavy damage to plant metabolism, reducing growth and yield. Injury to leaves due to excess ion-accumulation might be an important factor controlling the active size of the canopy (Francois and Maas, 1999). Increased salinity has an inverse relationship with stomatal conductance and net photosynthetic rate (Curtis and Läuchli, 1986; Lopez et al., 2002), leading to reduced photo-assimilation and dry matter production (Rozeff, 1995; Lingle and Weigand, 1997).
Salt tolerant plants adopt many strategies that range from morpho-anatomical to physiological and biochemical in
nature (Cheeseman, 1988; Zhu, 2001). The physiological ones include the exclusion of ions into physiologically less active parts (Schachtman and Munns, 1992), better selectivity of K+ over Na+ (Wilson et al., 2000), and synthesis of compatible osmotica for osmo-protection (Sakemoto and Murata, 2002). Plant tolerance to salinity may be more related to the Na+:K+ ratio in the cell than the absolute Na+ concentration (Benzyl and Reuveni, 1994; Qian et al., 2001). The salinity resistance in maize has been mainly related to greater flux and cytoplasmic K+ concentration (Hajibagheri et al., 1989). Reduction in water uptake by the root and hampered cell-water relations are both due to the osmotic component of salinity (Cheeseman, 1988; Wahid et al., 1999a). Tolerant plants adjust osmotically by the synthesis of highly water soluble compatible osmotica (e.g. glycinebetaine, free proline, and low molecular weight sugars) and maintain turgor. Among these, free proline ameliorates salt-induced oxidative damage to membranes (Jain et al., 2001), and glycinebetaine buffers the cellular redox potential (Hare et al., 1998), maintains Na+ balance between the cytoplasm and the vacuole (Subbarao et al., 2001), and protects cytoplasmic membranes during salt stress (Hare et al., 1998; Rehman et al., 2002; Sakamoto and Murata, 2002). Likewise, both reducing and non-reducing sugars contribute to turgor maintenance under salt or water stress (Cheeseman, 1988; Garg et al., 2002).
*E-mail: drawahid2001@yahoo.com