Bot. Bull. Acad. Sin. (2004) 45: 159-164
Zhang et al. Effects of C. laurentii in combination with SBC on biocontrol of postharvest green mold decay
Effects of Cryptococcus laurentii (Kufferath) Skinner in combination with sodium bicarbonate on biocontrol of postharvest green mold decay of citrus fruit
Hong-Yin Zhang1, Cheng-Xin Fu2, Xiao-Dong Zheng1,*, Dan He1, Li-Jun Shan1, and Xi Zhan1
1Department of Food Science and Nutrition, 2Department of Biology, Zhejiang University, Hangzhou 310029, People's Republic of China
(Received November 12, 2003; Accepted February 16, 2004)
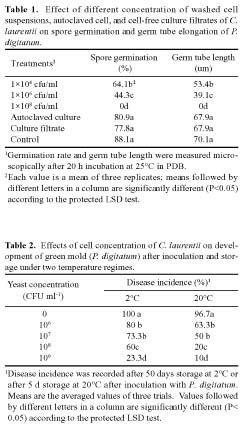
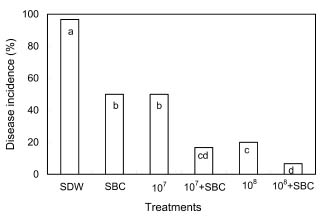
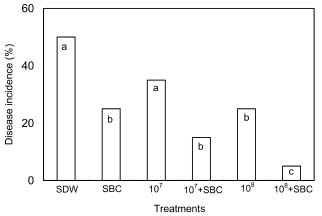
Abstract. The potential of using Cryptococcus laurentii alone or in combination with sodium bicarbonate solution for control of Penicillium digitatum (green mold) on oranges was investigated. Agar disks of C. laurentii NYDA cultures placed on PDA plates seeded with pathogen did not inhibit the growth of P. digitatum. Spore germination of P. digitatum in PDB was significantly controlled in the presence of living C. laurentii cell suspensions. Cryptococcus laurentii significantly controlled green mold on oranges after challenge with 5 × 104 spores/ml of P. digitatum. The higher the concentrations of the antagonist, the lower the disease incidence regardless of whether the fruit was stored at 20°C for 5 days or 2°C for 50 days. The efficacy of C. laurentii for control of green mold was improved when combined with sodium bicarbonate. The combination of sodium bicarbonate and C. laurentii could be an alternative to fungicides for control of postharvest green mold disease on citrus fruits.
Keywords: Biocontrol; Citrus fruits; Cryptococcus laurentii; Green mold; Sodium bicarbonate.
Introduction
Green mold of citrus, caused by Penicillium digitatum (Pers.:Fr.) Sacc., is one of the most economically important postharvest diseases of citrus worldwide (Smilanick et al., 1997; Caccioni et al., 1998; Holmes and Eckert, 1999; Palou et al., 2002). Currently, the disease is primarily controlled by applying synthetic fungicides such as imazalil and thiabendazole (Holmes and Eckert, 1999; Palou et al., 2001). However, alternative methods are needed because of concerns about environmental contamination and human health risks associated with fungicide residues (Wisniewski and Wilson, 1992) and because the widespread use of these chemicals in commercial packinghouses has led to the proliferation of resistant strains of the pathogens (Palou et al., 2002).
Microbial biocontrol agents have shown great potential as an alternative to synthetic fungicides for control of the postharvest decay of fruits and vegetables (Chalutz and Wilson, 1990; Wilson et al., 1991; Janisiewicz and Marchi, 1992; EI-Ghaouth et al., 2000). Strains of the yeast Cryptococcus laurentii (Kufferath) Skinner have been studied for the postharvest biological control of gray and blue mold rots of apples (Roberts, 1990a) and other fruits such as strawberries, kiwifruits, and table grapes (Lima et al., 1998), as well as for Mucor rot of pears (Roberts, 1990b).
Sodium bicarbonate (SBC) is another promising alternative to synthetic fungicides (Larrigaudière et al., 2002). SBC holds a generally recognized as safe (GRAS) classification by the United States Food and Drug Administration and is proposed to be exempt from residue tolerances on all agricultural commodities by the United States Environmental Protection Agency (Palou et al., 2001). It was previously reported that SBC salts reduce the development of blue and green mold decay in lemon, orange, and mandarin fruits (Smilanick et al., 1999; Palou et al., 2001, 2002).
Although both of the above-mentioned approaches have been shown to reduce postharvest rots of citrus, each exhibits limitations that can affect its commercial applicability. When used as independent treatments, none of these control methods has been clearly shown to offer a consistently economical level of disease control that would warrant its acceptance as a viable alternative to synthetic fungicides (Porat et al., 2002).
In the near future, the preferred alternative to synthetic fungicide treatments will probably be a combination of different methods (Teixidó et al., 2001). The objective of the present study is to evaluate the effects of combining SBC with the yeast biocontrol agent C. laurentii, in controlling green mold decay of citrus fruit.
Materials and Methods
Fruits
Oranges (Citrus sinensis [L.] Osbeck) cultivar `Bendizao' fruits were harvested at commercial maturity
*Corresponding author. Tel: 86-0571-86971167; Fax: 86-0571-86045315; E-mail: xdzheng@zju.edu.cn