Bot. Bull. Acad. Sin. (2004) 45: 197-202
Chen et al. Respiration and soluble sugars in methyl jasmonate-treated rice leaves
Changes in soluble sugar content and respiration rate in methyl jasmonate-treated rice leaves
Chien Teh CHEN1, Yen-Shuo SU1, and Ching Huei KAO2,*
1Department of Agricultural Chemistry, National Taiwan University, Taipei 106, Taiwan, Republic of China
2Department of Agronomy, National Taiwan University, Taipei 106, Taiwan, Republic of China
(Received December 8, 2003; Accepted March 4, 2004)
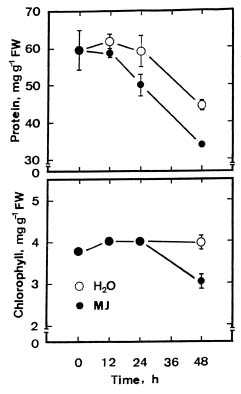
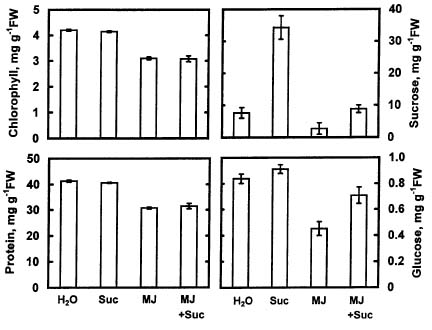
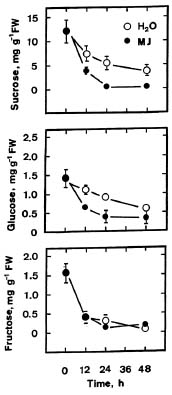
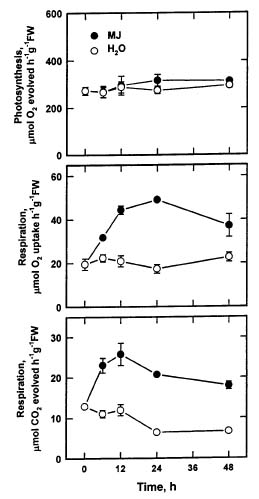
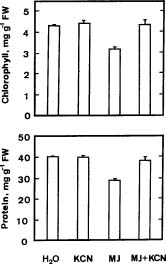
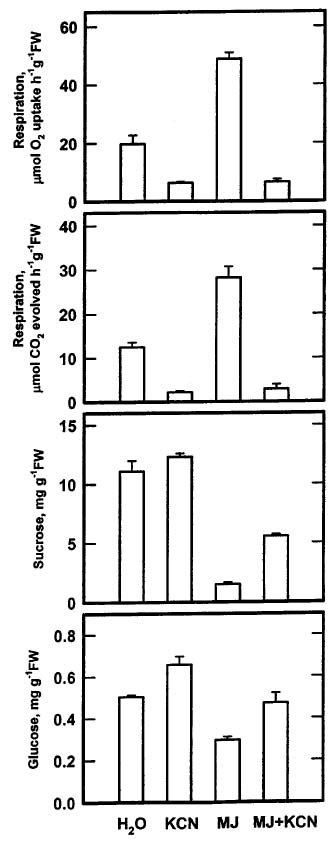
Abstract. The changes in respiration rate and soluble sugar content of detached rice (Oryza sativa) leaves were followed during methyl jasmonate (MJ)-promoted senescence. Increase in the respiration of detached rice leaves was observed as a consequence of methyl jasmonate (MJ) treatment for 48 h. However, MJ had no effect on photosynthetic rate in rice leaves. Sucrose and glucose contents, but not fructose content, were lower in MJ-treated rice leaves than in control leaves. KCN, an inhibitor of cytochrome c oxidase, was effective in inhibiting MJ-increased respiration rate, in reducing MJ-decreased sucrose and glucose contents, and in retarding MJ-promoted senescence of rice leaves. Exogenous addition of sucrose to MJ-treated rice leaves increased endogenous sucrose and glucose contents, but did not prevent MJ-promoted senescence of rice leaves. The role of respiration and sucrose in regulating MJ-promoted senescence of rice leaves is discussed.
Keywords: Fructose; Glucose; Methyl jasmonate; Oryza sativa; Respiration; Senescence; Sucrose.
Introduction
Jasmonic acid and its methyl ester, methyl jasmonate (MJ), have been proposed as naturally occurring plant growth regulators because of their wide natural distribution (Meyer et al., 1991) and their effects on many physiological processes in plants (Sembdner and Parthier, 1993). Jasmonates have been shown to be powerful promoters of leaf senescence (Ueda and Kato, 1981; Weidhase et al., 1987; Cuello et al., 1990). We also reported that MJ was effective in promoting the senescence of detached rice leaves in the dark (Chou and Kao, 1992; Tsai et al., 1996; Hung and Kao, 1997; Chen and Kao, 1998).
The relation between respiration and leaf senescence is not a simple one. If senescence were viewed as a general decline in function (Thimann, 1980), a steady decline of respiration rate would be expected. On the other hand, synthesis of degradative enzymes and nutrient mobilization processes during senescence all seem to require a higher respiration rate (Thimann, 1980). Studies on respiration changes (determined either by CO2 evolved or O2 uptake) during senescence have been made on either attached leaves or detached leaves of several plant species (Rhodes, 1980). In general, respiration rate increases during senescence in various plant materials. Kinetin, a synthetic cytokinin, at a concentration which retards senescence of detached oat leaves, completely prevents the respiration rise (Tetley and Thimann, 1974; Satler and
Thimann, 1983). Recent experiments by Dufour et al. (2000) demonstrated that the respiration function is a key factor contributing to a shorter lifespan for the filamentous fungus Podospura anserine.
Several reports show that the soluble sugar content often goes up, not down, in senescing leaves (Shiroya et al., 1961; Trippi, 1965; Egli et al., 1980; Lazan et al., 1983; Crafts-Brandner et al., 1984), and sugar accumulation in leaves suppresses the expression of some genes related to photosynthesis (Gan and Amansino, 1997; Jang et al., 1997). It has even been proposed that an elevated sugar content may actually cause senescence (Lazan et al., 1983).
MJ treatment has been shown to stimulate respiration of apple fruit (Miszczak et al., 1995; Fan and Mattheis, 1999), but it did not change the normal climacteric rise in the respiration of mango fruit (Gonazalez et al., 2000). However, to our knowledge, no one has studied the effect of MJ on the change in respiration and soluble sugars in leaves. The work reported here, therefore, explores the relations between respiration, soluble sugars, and the MJ-promoted senescence of rice leaves.
Materials and Methods
Rice (Oryza sativa cv. Taichung Native 1) seeds were sterilized with 2.5% sodium hypochlorite for 15 min and washed extensively with distilled water. These seeds were then germinated in Petri dishes with wetted filter paper at 37°C in the dark. After 48 h incubation, uniformly germinated seeds were selected and cultivated in a 500 ml beaker containing half-strength Kimura B solution as described previously (Chu and Lee, 1989). The hydroponi
*Corresponding author. Tel: 886-2-23698159; Fax: 886-2-23620879; E-mail: kaoch@ntu.edu.tw