Bot. Bull. Acad. Sin. (2004) 45: 203-212
Wang et al. Copper-induced change in antioxidation
Copper-induced stress and antioxidative responses in roots of Brassica juncea L.
Song-Hua Wang1,2, Zhi-Min Yang1,*, Hong Yang1, Bo Lu1, Shao-Qong Li1, and Ya-Ping Lu1
1Department of Biochemistry and Molecular Biology, College of Life Science, Nanjing Agricultural University, Nanjing, 210095, P.R. China
2Department of Applied Biology, Anhui Technical Teacher's College, Fengyang, Anhui, P.R. China
(Received November 10, 2003; Accepted February 25, 2004)
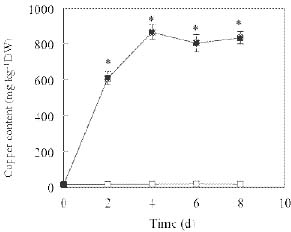
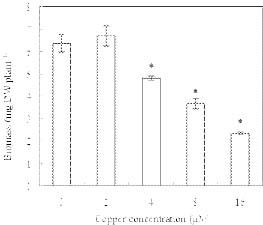
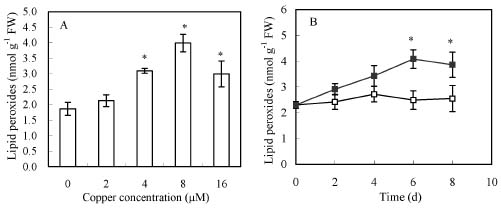
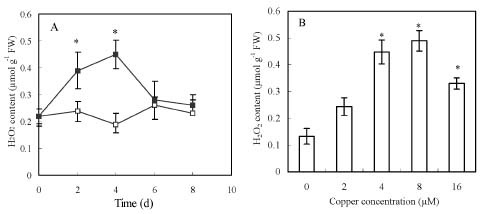
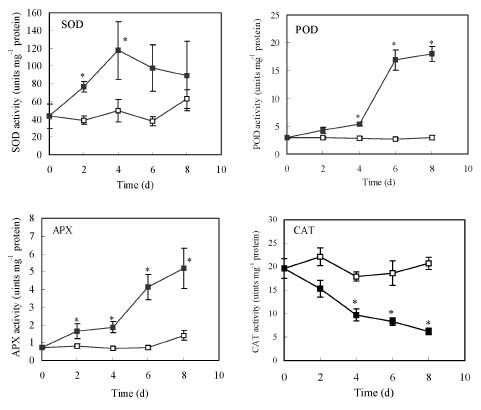
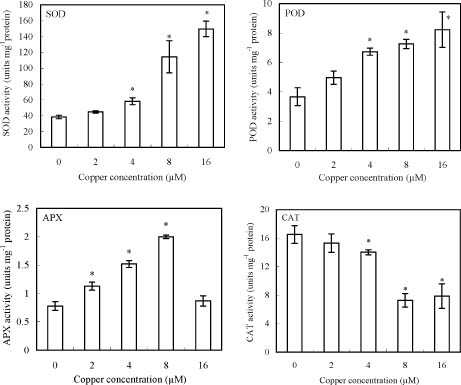
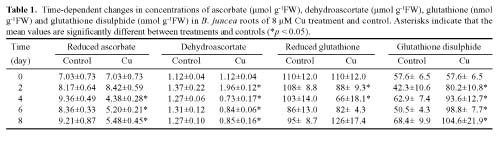
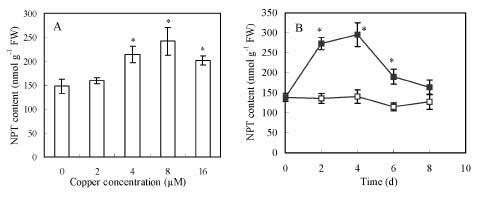
Abstract. Copper induced antioxidative reactions in the roots of Brassica juncea L. were investigated in both time-and concentration-dependent manners. The rapid uptake of Cu was observed immediately after the start of treatment. Application of Cu at 8 µM caused 50 percent reduction in biomass of Cu-treated roots as compared with control. Cu-induced root growth inhibition paralleled the level of root oxidative damage. Treatment with Cu at 8 µM induced a twofold increase in H2O2 content during the first 4 d, but it declined to the basal level thereafter. We also observed a twofold increase in superoxide dismutase activities with 8 µM Cu during the first 2 d. The stimulation lasted for 4 d and then gradually declined. Activities of both ascorbate peroxidase and guaiacol peroxidase in roots were found to be low during the first 4 d after seedling exposure to 8 µM Cu, but significantly increased after that, suggesting that increased enzyme activities would be responsible for the removal of H2O2. Catalase activities were always suppressed under Cu stress. Treatment of seedlings with 8 µM Cu induced general decreases in both reduced ascorbate and dehydroascorbate. The reduced glutathione content decreased at early stages of Cu treatment. However, it was restored to the level of controls thereafter. In contrast, the oxidized glutathione contents showed a progressive increase during the time of Cu treatment. The total non-protein thiol content was shown to increase during the first several days, but it declined at later stages.
Keywords: Antioxidative enzymes; Brassica juncea L.; Copper; Hydrogen peroxide; Non-protein thiols.
Introduction
Copper, an essential micronutrient, plays a vital role in maintaining normal metabolism in higher plants. It is involved in a wide range of biochemical and physiological processes. For example, Cu is required as a cofactor of Cu-Zn superoxide dismutase (SOD, EC 1.1.5.1.1) (Bowler et al., 1992). It also participates in electron-transfer reactions of photosynthesis in the form of plastocyanin (Raven et al., 1999). However, Cu at high levels becomes strongly phytotoxic to cells and causes inhibition of plant growth or even death (Mocquot et al., 1996; Weckx and Clijsters, 1996; Chen et al., 2000). Studies from some plant species demonstrate that excess Cu in plant growth medium induces formation of reactive oxygen species in treated-tissues (Cuyers et al., 2000; Groppa et al., 2001). Cu-induced generation of hydrogen peroxide, hydroxyl redicals, or other reactive oxygen species (ROS) has been directly correlated with the damage to proteins and lipids (De Vos et al., 1991; Murphy and Taiz, 1997). Photosynthesis is also sensitive to excessive Cu, and the pigment and protein components of photosynthetic membranes are the targets
(Pätsikkä et al., 2002). In addition, Cu toxicity is related to disturbances in the uptake of other essential elements (van Assche and Clijsters, 1990; Pätsikkä et al., 2002).
The phenomenon of Cu activating the production of ROS is described as oxidative stress (Luna et al., 1994). To repair the oxidative damage initiated by ROS, some tolerant plants have established protective mechanisms. The primary constituents of these protective mechanisms include two scavenging systems: enzymatic and non-enzymatic. Enzymatic scavengers, such as superoxide dismutase, are involved in the detoxification of O2.-; peroxidase, catalase, and enzymes of the ascorbate-glutathione cycle are related to the removal of H2O2 (De Vos et al., 1991; Murphy and Taiz, 1997); and non-enzymatic redicals-scavengers such as ascorbate and glutathione have been interpreted as the key antioxidants for the removal of H2O2 in plant cells, thus reducing the accumulation of the free radicals (van Assche and Clijsters, 1990; Foyer, 1993; Gupta et al., 1999). Evidence from several plant species reveals that Cu caused oxidative stress by mediating the activities of antioxidative enzymes (Luna et al., 1994; Gupta et al., 1999; Cuyers et al., 2000). Savouré et al. (1999) reported that Cu treatment triggered distinct oxidative defense mechanisms in Nicotiana plumbaginifolia. However, contrasting results are present. For example, in leaves of Cu-exposed Avena sativa plants, superoxide
*Corresponding author. Tel: 86-25-84395057; Fax: 86-25-84073096; E-mail: zmyang@njau.edu.cn