Bot. Bull. Acad. Sin. (2004) 45: 213-220
Deng et al. Gas exchange characteristics of an invasive weed species and its indigenous congener
Gas exchange characteristics of the invasive species Mikania micrantha and its indigenous congener M. cordata (Asteraceae) in South China
Xiong DENG1,2, Wan-Hui YE1,*, Hui-Ling FENG1, Qi-He YANG1, Hong-Ling CAO1, Kai-yang XU1, and Yun ZHANG1
1South China Botanic Garden, Chinese Academy of Sciences, Guangzhou 510650, P.R. China
2South China Institute of Environmental Sciences, State Environmental Protection Administration of China, Guangzhou 510650, P.R. China
(Received November 5, 2003; Accepted March 29, 2004)
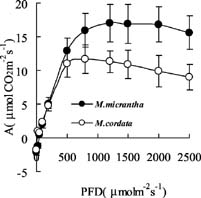
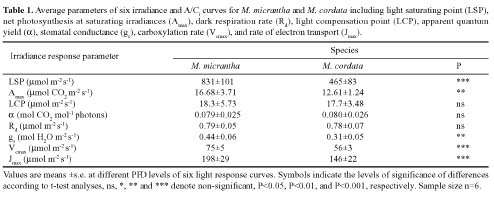
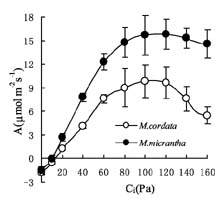
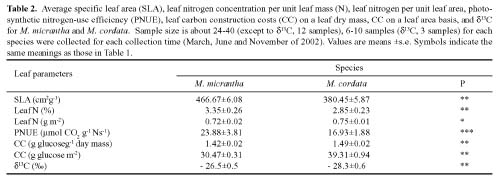
Abstract. In South China, the photosynthetic characteristics of an invasive exotic weed Mikania micrantha H.B. Kunth and its indigenous close congener Mikania cordata (Burm. f.) B.L. Robinson were studied. Mikania micrantha exhibited a greater biochemical capacity for photosynthesis than M. cordata, as the former had higher Vcmax and Jmax, which lead to its higher maximum assimilation rate (Amax) values. Similar light compensation point, apparent quantum yield (a), and dark respiration (Rd), suggested a similar shade-tolerance. Higher Amax, light saturating point, and CO2 saturating point give M. micrantha greater potential to acclimate to brighter environments and a larger pool of available carbon. These differences in photosynthetic characteristics could be a basis for their niche partitioning. The leaf traits of the native and invasive Mikania species were as follows: higher construction cost (CC), lower specific leaf area (SLA), as well as lower mass-based leaf N and photosynthetic nitrogen-use efficiency (PNUE) for the native species, but M. micrantha had lower area-based leaf N. The higher water use efficiency (WUE) and d13C observed in M. micrantha was probably due to its high photosynthetic capacity and not to reduced stomatal conductance, explaining the absence of any trade-off between WUE and PNUE.
Keywords: Invasive species, Mikania; Nitrogen use efficiency; Photosynthetic characteristics; Specific leaf area.
Introduction
The impact of invasive weeds on native species, communities, and ecosystems has been widely recognized for decades (Elton, 1958; Lodge, 1993; Simberloff, 1997; Vitousek et al., 1997; Mack et al., 2000). However, we still lack a fundamental understanding of the mechanisms by which invasive plants succeed, an understanding that may eventually prove predictive and useful for control efforts (Baruch and Goldstein, 1999; Mack et al., 2000; Kolar and Lodge, 2001). Comparison of closely related invasive and non-invasive congeners that share morphological and life-history traits is an effective approach to identifying mechanisms of invasive plant success (Schierenbeck and Marshall, 1993; Mack, 1996). Distinguishing the ecophysiological traits that confer invasion success (Mack, 1996; Xu et al., 2004) is more precise than working from the concept of an "ideal weed" (Baker, 1965, 1974), which summarizes common attributes of plant invaders from broad surveys (David and Baruch, 2000; Mack, 1996). This latter approach has been widely criticized for its inability to predict the outcome of any specific introduction and for the numerous exceptions to any unique suite of traits ex
pressed by plant invaders (Roy, 1990; Rejmánek, 1995; Rejmánek and Richardson, 1996).
Mikania micrantha H.B. Kunth, commonly known as mile-a-minute, is an extremely fast growing, sprawling, perennial vine and one of the world's most notorious invaders (Holm et al., 1977; Cronk and Fuller, 1995). With its rapid growth, ready rooting at nodes, smothering habit, and prolific seed production, M. micrantha rapidly colonizes disturbed habitats, retarding the growth of crops or natural vegetation by competing and producing plant inhibitors. In addition to its native distribution in tropical America, it has spread to Mauritus, India, Sri Lanka, Bangladesh, Southeast Asia, and the Pacific. Mikania cordata (Burm. f.) B. L. Robinson is the native congener of M. micrantha. They appear to be similar species, as they share similar morphologies and life histories, and their identities have been confused in some documents (Holm et al., 1977; Kong et al., 2000). However, in sharp contrast to M. micrantha, M. cordata grows slowly and has not been found to be harmful to native plants or habitats in southern subtropical China (Kong et al., 2000). Additionally, while M. micrantha can thrive in sunny environments, M. cordata can only be found in relatively shady and humid wetlands, and it is more drought-tolerant than (Huang et al., 2000).
It is interesting that M. micrantha is not even considered a weed in its native Brazil, but now is a notorious
*Corresponding author. Tel: (+86) 13710546160; E-mail: why@scib.ac.cn