Bot. Bull. Acad. Sin. (2004) 45: 221-228
Park Allozyme variation of Far East Euphorbia
Comparisons of allozyme variation of narrow endemic and widespread species of Far East Euphorbia (Euphorbiaceae)
Ki-Ryong PARK*
Department of Biology, Kyung-Nam University, Masan 631-701, Korea
(Received January 2, 2004; Accepted April 23, 2004)
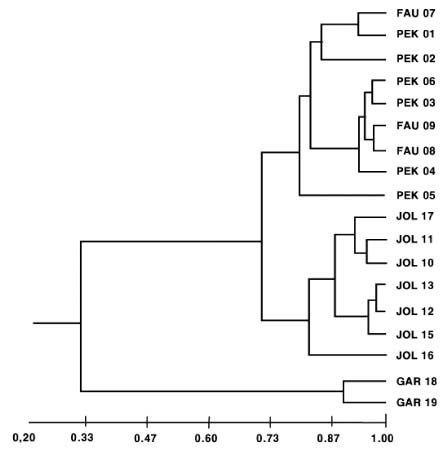
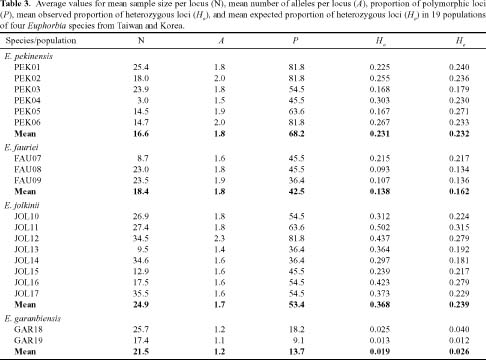
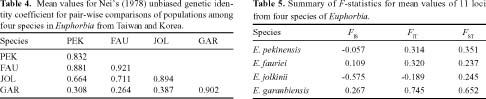
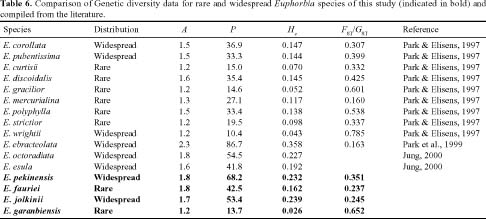
Abstract. Genetic variation from 19 populations representing four species of Euphorbia was compared using isozyme markers. Euphorbia fauriei and E. garanbiensis are rare endemic species in Korea and Taiwan, respectively while E. pekinensis and E. jolkinii are widespread in Korea and Taiwan. A low level of genetic variation and high genetic differentiation among populations was found in E. garanbiensis, which has a restricted distribution, small population sizes, gravity-dispersed seeds, and is possibly a self-pollinator. Alternatively, E. garanbiensis may have recently evolved from a continental progenitor or may be a relictual lineage from the continental flora. The high genetic diversity found in E. fauriei is unusual and may be explained by its recent origin from a widespread continental progenitor, E. pekinensis. An alternative explanation is that E. fauriei survived in refugia during the last glaciation in the Korean peninsula. The idea of progenitor-derivative species pairs between E. pekinensis and E. fauriei is supported by the high genetic identity values between the two species and the putative derivative E. fauriei having no unique allele and lacking some rare alleles present in the putative progenitor E. pekinensis.
Keywords: Allozyme variation; Euphorbia fauriei; Euphorbia garanbiensis; Euphorbia jolkinii; Euphorbia pekinensis.
Introduction
Most rare and endemic plant species have low levels of genetic variability due to the effects of small population sizes (Hamrick and Godt, 1989; Ellstrand and Elam, 1993; Gitzendanner and Soltis, 2000; Smith and Pham, 1996; Sherman-Broyles et al., 1992). When compared with wide-ranging taxa, rare species are heavily impacted by genetic drift, the founder effect, and directional selection with high levels of inbreeding decreasing genetic diversity by eliminating polymorphic loci and reducing alleles per polymorphic locus (Karron, 1987; Barrett and Kohn, 1991; Sherman-Broyles et al., 1992; Dodd and Helenurm, 2002; Gitzendanner and Soltis, 2000). However, recent comparative studies of genetic variation between rare and widespread species have demonstrated that several rare species were as polymorphic as their widespread congeners (Dodd and Helenurm, 2002; Gitzendanner and Soltis, 2000). Thus, it is difficult to state that species with small populations and limited geographic range always have low genetic diversity. A common approach to determining the cause of differences in genetic variation between rare and widespread species has been to compare phylogenetically related species pairs (Edwards and Wyatt, 1994; Dodd and Helenurm, 2002; Gitzendanner and Soltis, 2000).
Euphorbia jolkinii, E. pekinensis, and E. fauriei are herbaceous perennial species of Far Eastern Euphorbia subg. Esula. Phylogenetic studies of these species using morphological and chemical data found them to be closely related and to form a distinct clade (Park et al., 1999a). The
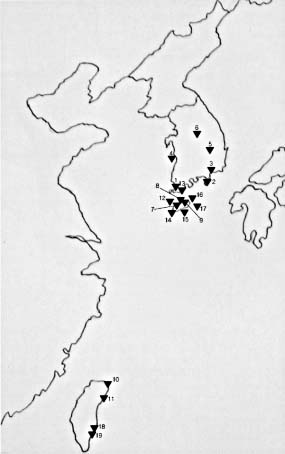
Korean endemic E. fauriei is a rare plant with a narrow distribution on Mt. Halla of Jeju Island and seemingly restricted to the open, rocky areas of the mountaintop (ca. 1,500 m). This species is diploid (2n = 28), insect pollinated, and perennial, growing to 10-20 cm in height. Euphorbia pekinensis is morphologically most similar to E. fauriei. It is a more widespread and morphologically variable species in Korea, Japan, and northeast China. Despite a high degree of morphological and cytological similarity between the two species, E. fauriei is well distinguished from E. pekinensis by several quantitative characters (Park et al., 2002). Recent cladistic analyses of morphological data indicate that these two species form a sister-group relationship (Park et al., 1999a). Euphorbia jolkinii, a sister group of E. pekinensis + E. fauriei, is a salt-tolerant species inhabiting rocky areas along the seacoasts of Korea, Japan, and Taiwan (Lin and Hsieh, 1991).
Euphorbia garanbiensis, a member of Euphorbia subg. Chamaesyce, is a rare endemic species of the Oluanpi Peninsula, the southernmost tip of Taiwan (Lin et al., 1991). It inhabits sandy areas and sea-cliffs along the coast with known populations restricted to a few localities in Pingtung County Taiwan (Lin et al., 1991). This species is a perennial, distinguished from the other three species by its prostrate habit and bearing conspicuous appendages on the rim of the cyathium. There have been few studies of the reproductive biology in Euphorbia, although self-pollination has been reported in several weedy species of the subg. Chamaesyce (Krombein, 1961; Ehrenfeld, 1976).
In the genus Euphorbia, the structure of the cyathium consists of four or five nectar-producing glands that have been considered an adaptation to insect pollination (Cronquist, 1968). A diverse array of insects, such as ants,
*Corresponding author. E-mail: park@kyungnam.ac.kr