Bot. Bull. Acad. Sin. (2004) 45: 229-236
Guan et al. Plasticity in Mosla chinensis and M. scabra
Phenotypic plasticity of Mosla chinensis and M. scabra (Labiatae) response to soil water status
Bao-Hua GUAN, Ying GE, and Jie CHANG*
Zhejiang University, College of Life Science, Xixi Campus, 232 Wensan Road, Hangzhou 310012, P. R. China
(Received October 8, 2003; Accepted April 1, 2004)
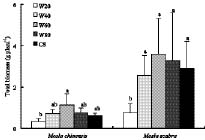
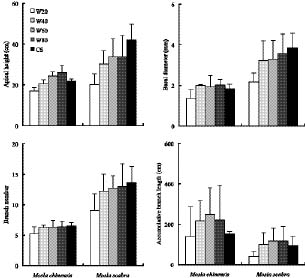
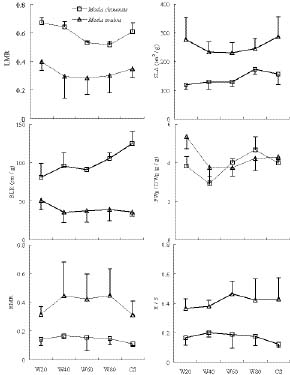
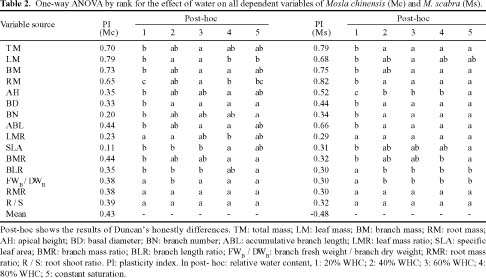
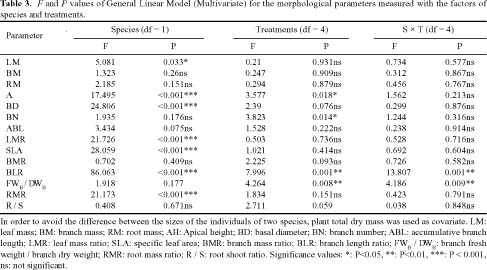
Abstract. The growth and architectural plasticity of Mosla chinensis Maxim. in response to soil water status were compared with the congeneric plant, Mosla scabra (Thunb.) C. Y. Wu et H. W. Li. Two-week-old seedlings were exposed to five levels of soil water for a 6-week period. The results indicated that: an individual's total mass, root mass, apical height, basal diameter, accumulative branch length and branch fresh weight / dry weight ratio (FWB / DWB) of both species had high plasticity in response to soil water content (P < 0.05), and the plasticity of these traits in M. scabra is mostly higher than in M. chinensis. The leaf mass ratio (LMR), specific leaf area (SLA), root mass ratio (RMR), and root / shoot ratio (R / S) of both species had low plasticity. Furthermore, leaf mass, branch mass, branch mass ratio (BMR), and branch length ratio (BLR) had high plasticity (P < 0.05) in M. chinensis but not in M. scabra (P > 0.05) while branch number exhibited contrary trends. In response to soil water, M. scabra adjusted the traits of total mass and size, in terms of a bigger PI, more than M. chinensis while M. chinensis only adjusted partial branch and root traits, such as BMR, BLR, FWB / DWB, RMR and R / S, more than M. scabra. The optimum water niches (OWN) of both M. chinensis and M. scabra are from 40% soil water holding capacity (WHC) to constant saturation, but M. chinensis is only found in relatively dry environments while M. scabra is distributed from dry to wet environments in the field, so the actual water niche (AWN) was separated from the OWN in M. chinensis, but not in M. scabra. Mosla chinensis grew slower and remained smaller than M. scabra and other neighbor species in the field, and it therefore had no competitive superiority in the community. Mosla scabra was very competitive because of its higher yield and taller growth.
Keywords: Interspecific difference; Mosla chinensis; Mosla scabra; Phenotypic plasticity; Relative soil water content; Water niche.
Introduction
Plants of the same genotype can produce different phenotypes in different environments, a property called phenotypic plasticity (Sultan, 2001). Adaptive phenotypic plasticity is the predicted evolutionary response to environmental factors, such as soil moisture in plant habitats (Sultan and Bazzaz, 1993). Differences among species in plasticity patterns may contribute to their differences in ecological breadth with respect to soil factors and climatic conditions (Cook et al., 1980). Bell and Sultan (1999) investigated how two Polygonum species altered their root growth and deployment over time in response to different soil moisture conditions, and they found that species differences in plastic response to specific moisture conditions would correspond to differences in their field distribution. Ryser and Eek (2000) reported two congeneric grass species with contrasting shade tolerance responded to low resource availability, and they indicated that interspecific differences in phenotypic plasticity were crucial factors for survival and distribution.
It is well known that the distribution and abundance of most plant species greatly depend on water supply
(Schulze et al., 1987). The influence of water availability on plant performance in natural and managed ecosystems has been of great concern in plant physiological ecology and related disciplines (Schulze et al., 1987). Plant growth may decrease in dry soils due to tissue dehydration as well as reduced mineral availability (Fitter and Hay, 1993).Conversely, flooding also reduces plant growth by decreasing the availability of oxygen to roots (Etherington, 1984; Ernst, 1990). There have been a number of studies on the phenotypic plasticity of plants in response to the water supply (Stevens et al., 1997; rùtek, 1997; Bell and Sultan, 1999; Clifton-Brown and Lewandowski, 2000; Kotowski et al., 2001), including a lot of studies elaborating the effect of plasticity on local traits for individuals, e.g., leaf and root traits (Bell and Sultan, 1999; Clifton-Brown and Lewandowski, 2000; Ryser and Eek, 2000).
Mosla chinensis Maxim., distributed in east, south, middle and southwest China (Fang et al., 1986), is a medicinal herb which contains many volatile oils and has a long history of use in China for rheum and heliosis (Zhang, 1989; Fang et al., 1986). Many studies about the medicinal components and physiology of M. chinensis have appeared (Zhang and Xu, 1988; Zheng et al., 1996; Zhou et al., 1996, 1998; Pan et al., 1997), but ecological studies of it are rare (Ge et al., 1999; Ge and Chang, 2001; Guan et al., 2003). Though its distribution in the field is wide, M. chinensis has only a few individuals in each population,
*Corresponding author. Tel & Fax: +86 571 8797 2193; E-mail: jchang@mail.hz.zj.cn