Bot. Bull. Acad. Sin. (2004) 45: 237-245
So The occurrence of extrafloral nectaries in Hong Kong plants
The occurrence of extrafloral nectaries in Hong Kong plants
May Ling SO*
Biology Department, Hong Kong Baptist University, 224 Waterloo Road, Hong Kong
(Received September 3, 2003; Accepted March 2, 2004)
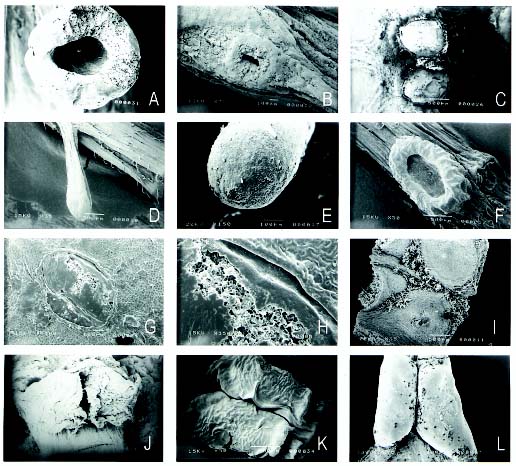
Abstract. This is a study of the extrafloral nectaries in Hong Kong plants. Five major types can be discerned: button-shaped, cup-shaped, stalk-shaped, pit-shaped, and pore-shaped. Euphorbiaceae is the largest family with extrafloral nectaries which are always visible structures, attracting ants. SEM micrographs of extrafloral nectaries are included.
Keywords: Extrafloral nectaries; Hong Kong plants.
Introduction
Nectaries not involved in pollination are called extrafloral nectaries (EFNs), sugar producing glands found outside the flower. They occur in at least 66 families (Elias, 1983), many in the tropics. Hong Kong, situated in a subtropical region, is rich in flowering plants, many of them either cultivated in parks and gardens or grow in the wild in many countryparks. EFNs have been shown to play an important part in the defense mechanism of plants against herbivores (Nees, 2003). Even though the debate over the role of extrafloral nectaries has lasted over a century, numerous scientists are still attracted by the role these small, fascinating structures play in the life of a flowering plant (Bentley, 1977; Koptur, 1992). The animals usually associated with EFNs are ants, and it is not uncommon to see ants crawling over leaves and petioles, looking for nectar. This ants/plant facultative mutualistic association has been described as beneficial to both parties (Pemberton, 1998). The ants, while obtaining food for their colony, can deter other herbivores from attacking the plants. Heil et al. (2001) even showed that the production of extrafloral nectar in Macaranga tanarius is an induced, indirect defensive reponse that strongly reduces herbivory. In a series of experiments to investigate the various factors affecting nectar production in Macaranga tanarius (Heil et al., 2000), nectar production is highest in unfolded young leaves, and the rate of secretion remains relatively constant throughout the day, peaking at dusk. The composition of the secretion is mostly fructose, glucose, sucrose (Heil et al., 2000), with traces of various amino acids (Ness, 2003). The ontogeny of EFNs in Cappris retusa has been studied by Di Sapio et al. (2001), who confirmed that nectar secretion appears early in the development process. Studies by Farji
Brenar et al. (1992) also indicate that secretion is apparently related to ant patrolling activities. Ness (2003) also showed that production of nectar increases two- to three-fold when leaves are attacked by caterpillars, together with further attraction of ant bodyguards. The presence of EFNs may further add an ecological advantage to these plants in self-protection, reduce vegetative damage, and help to prevent heavy foraging by other animals (Bentley, 1976; Pemberton, 1998). An experiment conducted on Sapium sebiferum by Rogers et al. (2003) indicated that simulated leaf herbivory significantly stimulated effluent production on EFN glands on seedlings. The beneficial effect of ants on the reproductive success of Dyckia floribunda (Bromeliaceae), an extrafloral nectary plant, is shown by Vesprini et al. (2003) that total seed production per plant was strongly affected by ant exclusion. However, other studies by Freitas et al. (2000) indicated no significant differences in either the degree of herbivory or in the reproductive output between stems of Croton sarcopetulus with ants and those without. Another hypothesis on the secretion of extrafloral nectar is that it is an attempt to distract insects from flowers (Rosenzweig, 2002). Since the secretion of nectar is an energy intensive process, Rosenzweig (2002) theorized that the cost of each extrafloral nectary divided by the cost of each flower must be less than the proportion of reproduction threatened by insect visits.
The purpose of this paper is to study the EFNs in Hong Kong plants and examine the type, shape, number, and position on the leaves. Extrafloral nectaries differ considerably in gross morphology. Since many of the EFNs are minute and inconspicuous, e.g. Acacia sp., they are easily overlooked.
Materials and Methods
A number of field trips to numerous localities in various parts of Hong Kong were conducted to look for plants
*Corresponding author. E-mail: mlso@hkbu.edu.hk