Bot. Bull. Acad. Sin. (2004) 45: 247-257
Chen and Chen HSPs of thermophilic and thermotolerant fungi from Taiwan
Heat shock proteins of thermophilic and thermotolerant fungi from Taiwan
Kuei-Yu CHEN1,* and Zuei-Ching CHEN2
1Department of Biology, Chinese Culture University, Taipei 111, Taiwan
2Department of Botany, National Taiwan University, Taipei 106, Taiwan
(Received December 18, 2003; Accepted April 22, 2004)
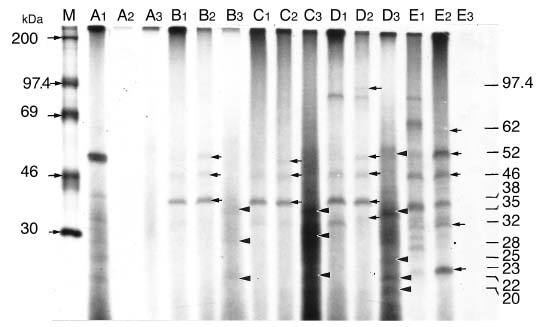
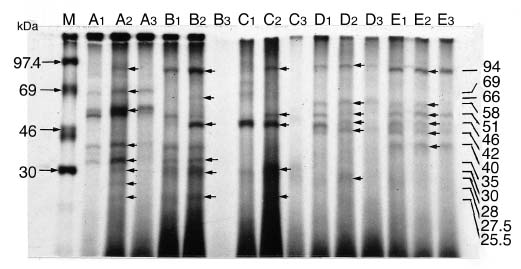
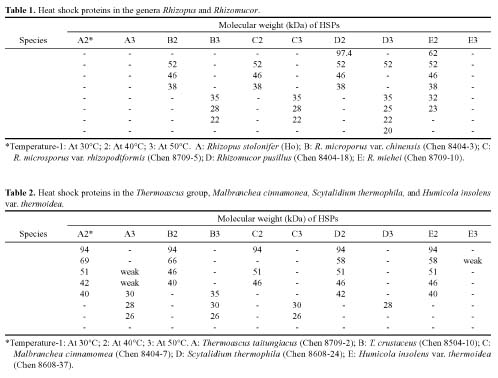
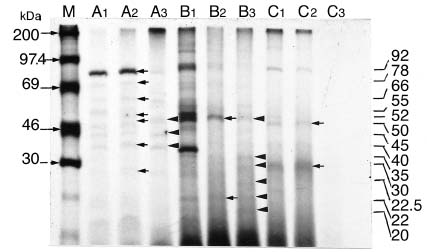
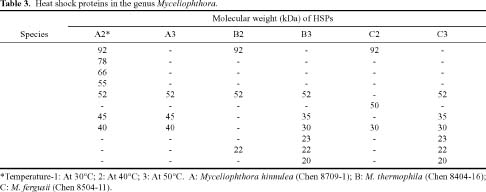
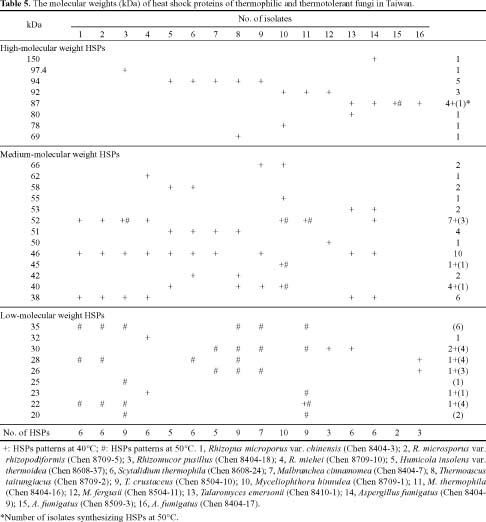
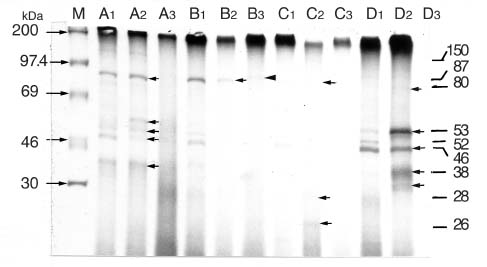
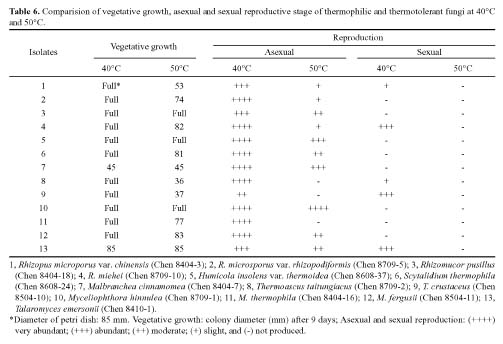
Abstract. Fifteen species of thermophilic and four species of thermotolerant fungi were isolated from soil samples collected at various localities in Taiwan. Fourteen species and three strains responded to three h of thermal stress at elevated temperature from 30°C to 50°C by synthesizing 30 heat shock proteins (HSPs) with molecular weights ranging between 20-150 kDa. Heat shock treatments at 40°C, resulted in the synthesis of 22 HSPs with molecular weights of 30-150 kDa. Nine of the seventeen fungal species produced a 46 kDa HSP, seven species a 52 kDa HSP, and five species a 94 kDa HSP. Four of the seventeen species produced two kinds of HSPs with molecular weights of 87 kDa and 40 kDa. Ten fungal strains responded to an elevated temperature of 50°C and synthesized 12 HSPs of molecular weights ranging from 20 to 92 kDa. Six species produced a 35 kDa HSP, and four species produced HSPs with molecular weights of 30 kDa, 28 kDa and 22 kDa. When subjected to heat shock treatments at 40°C, thermophilic and thermotolerant fungi synthesized mostly high and medium molecular weight HSPs while at 50°C, they synthesized mostly low molecular weight HSPs.
Keywords: Heat shock proteins; Thermophilic fungi; Thermotolerant fungi.
Introduction
Fungi are heterotrophic eukaryotes which exhibit a great diversity in morphology and distribution. Some species can survive at the elevated temperatures of hot springs, in the high salinity of sea-water, or in various other adverse environments. Environmental stresses or stimuli will change the structure or metabolism of an organism by acting as elicitors which affect gene expression and result in the synthesis of stress-specific compounds to protect the organism.
Adverse factors can be either biotic or abiotic. Biotic factors include bacteria, fungi, insects, or disease-causing organisms. They elicit changes in host genetic expression so that stress-specific compounds are synthesized to enhance host resistance to the foreign organism. Abiotic factors include temperature, excess water (Ben-Zioni et al., 1967; Hsiao, 1970), salinity (Ben-Zioni et al., 1967; LaRosa et al., 1989; Burk and Jennings, 1990), anaerobic conditions (Frelling and Bennett, 1985), heavy metals (Jackson et al., 1984; Curle and Kapoor, 1988; Gruhn and Miller, 1991), growth regulator (Heikkila et al., 1984), ultraviolet irradiation (Chappell and Hahlbrock, 1984), metabolic repressors, oxidizers (Curle and Kapoor, 1988), famine (Curle and Kapoor, 1988), and pH (LeJohn and Braithwaite, 1984). Among the environmental stressors listed above, thermal stress has been most widely studied. Both heat shock and cold shock can induce the synthesis or storage of a group
of proteins which increase resistance to thermal stress (Ketola-Pirie and Atkinson, 1983; Yacoob and Filion, 1987).
When thermal stress is applied, the most prominent physiological reactions are the production of a set of novel proteins or an increase in the quantity of certain types of existing proteins. These proteins are known as heat shock proteins (HSPs) (Ketola-Pirie and Atkinson, 1983; Neidhardt et al., 1984; Lindquist, 1986; Lindquist and Craig, 1988; Freeman et al., 1989). This phenomenon was first discovered in 1962 by Ritossa in fruit fly larvae (Drosophila buskii) and has since been manifested in other living organisms (Lindquist, 1986; Lindquist and Craig, 1988).
The optimum temperature for the production of HSPs varies from organism to organism. The heat shock temperature range for Escherichia coli is 43- 47°C (Neidhart et al., 1984), for the yeast Saccharomyces cerevisiae 36°C (McAlister and Finkelstein, 1980), and for the sickle fungus Fusarium oxysporum 40°C or 43°C (Freeman et al., 1989). In general, a rise of 5°C above the normal physiological temperature will induce the synthesis of HSPs.
Heat shock proteins can be classified into three categories according to their molecular size: (I) high-molecular size, with molecular weight between 69 and 120 kDa, (II) medium-molecular size, with molecular weight between 39 and 68 kDa, and (III) low-molecular size, with molecular weight below 38 kDa.
In fungi, the synthesis of HSPs is a rapid process. For example, 10 min after a heat treatment, F. oxysporum began to synthesize HSPs (Freeman et al., 1989) while for S. cerevisiae, a period of 20 to 30 min was required. The synthesis of HSPs peaked at 60 min after heat treatment of
*Corresponding author. Tel: 886-2-28610511-652; Fax: 886-2-28623724; E-mail: thermophilic@ms52.url.com.tw