Bot. Bull. Acad. Sin. (2004) 45: 267-274
Charng et al. NLS-transposase triggers high transposition efficiency
Fusion of the transposase with a classical nuclear localization signal to increase the transposition efficiency of Ac transposon
Yuh-Chyang CHARNG1,*, Hui-Ping LI1, Hung-Chun CHANG3, Kuan-Te LI1, Tzong-Hsiung HSEU2, and Jenn TU3
1Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
2Department of Life Science, National Tsing Hua University, Taiwan, Republic of China
3Institute of Botany, Academia Sinica, Taipei, Taiwan, Republic of China
(Received January 29, 2004; Accepted June 29, 2004)
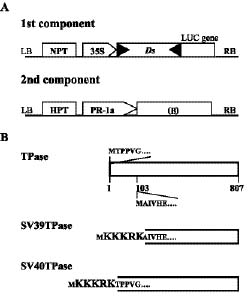
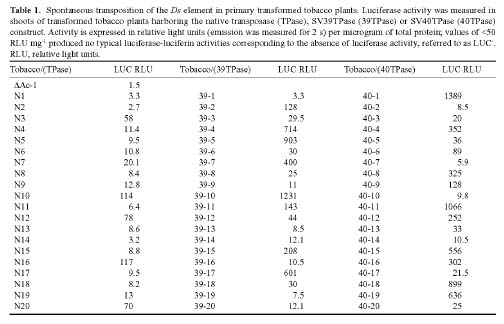
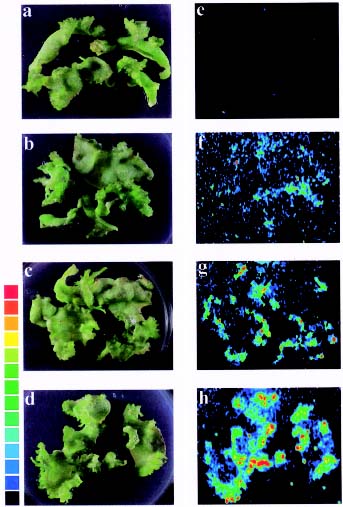
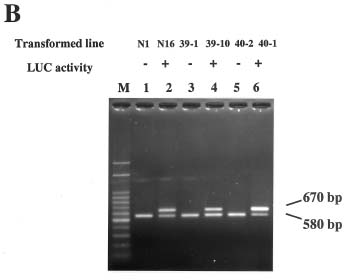
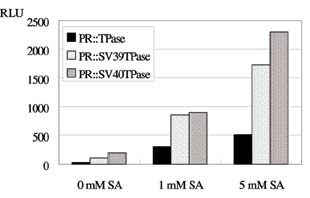
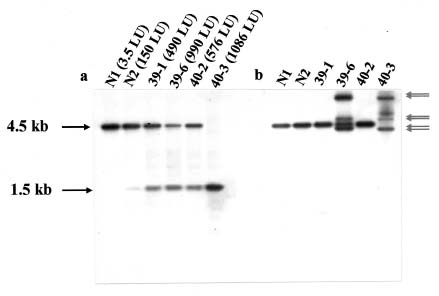
Abstract. A new strategy was applied to improve the transposition efficiency of the maize transposon Activator (Ac) in heterologous plants. The Ac transposase was fused with a classical nuclear localization signal (NLS) of SV40 to promote the transport of transposase into a nucleus. Base on this, two NLS-TPase constructs were yielded, one containing the full length transposase gene (termed as SV40TPase), the other containing the truncated transposase gene (lacking its first NLS-like signal, termed as SV39TPase). These two NLS-TPase genes were expressed in transgenic tobacco plants under the control of PR-1a promoter. Excision of non-autonomous transposable element (Ds) from luciferase (LUC) reporter gene constructs was employed to analyze the induction of Ac transposase containing NLS. Applying the LUC assay and PCR analysis, these new NLS-TPase sources triggered higher Ds excision efficiencies then the native transposase. Furthermore, the SV40TPase showed more ability then the SV39TPase to trigger the Ds element. The usage of this new inducible transposon for plant functional genomics is discussed.
Keywords: Ac transposase; Inducible transposon; Luciferase reporter gene; Nuclear localization signal.
Abbreviations: HPT, hygromycin phosphotransferase; NLS, Nuclear localization signal; NPT, neomycin phosphotransferase; LUC, luciferase; SA, salicylic acid; TPase, transposase.
Introduction
The maize transposon Activator (Ac) is an autonomous transposable element with a size of 4565 bp. It codes for a single gene product, the transposase (TPase), which together with the inverted repeats and about 300 bp from each end of the transposon and putative host factors is the only prerequisite for transposition of the Ac element in plants. The Ac element is active in a wide range of plant species, including several members of the Solanaceae, and in rice, carrot and Arabidopsis (Becker et al., 1986; Van Sluys et al., 1987; Knapp et al., 1988; Yoder et al., 1988; Houba-Herin et al., 1990; Izawa et al., 1991), and has proven to be a powerful genetic tool for yielding knockout mutants for plant functional genomic studies (for a review, see Haring et al., 1991). Recently, the International Rice Genome Sequencing Project (IRGSP) completed its sequencing of the entire rice (Oryza sativa) genome. Various strategies, including transposon tagging, have been used to produce a large population of mutant plants adequately assigning function to the abundant sequence information (for review see Jeon and An, 2001). However, using transposon as a tool to create knockout mutants in plants with large genomes, seems to require increased
transposition efficiency. To this end, Scofield et al. have fused the Ac TPase with the cauliflower mosaic virus (CaMV) 35S RNA promoter and found, in tobacco, no direct proportionality between the amounts of TPase mRNA and Ac/Ds transposition activity (Scofield et al., 1992). Furthermore, transpositions occur only at TPase transcription level, below a critical threshold (Scofield et al., 1993). One possible explanation is that above this threshold the TPase starts to aggregate and transpositions cease (Heinlein et al., 1994), forcing us to consider another approach to improving transposition efficiency.
In principle, in order to perform the transposition events, the TPase can be transported into the nuclei, a process mediated by specific signals called nuclear localization sequences (NLSs) (Stochaj and Silver, 1992a). The TPase has three NLSs near its amino-terminal end, NLS (44-62), NLS (159-178), and NLS (174-206), each of which is sufficient to direct GUS to the nucleus (Boehm et al., 1995). However, NLS (44-62) and NLS (159-178) are bipartite NLSs while the structure of NLS (174-206) is not in one of the three major NLS categories. Interestingly, all three sequences were determined to be "weak" NLSs or NLS-like signals (Heinlein et al., 1994; Wang et al., 1998 and personal communication). We predict then that fusing a classical-NLS sequence (e.g. a source from SV40) to the TPase protein could promote the transport of the Ac TPase and consequently increase transposition efficiency.
*Corresponding author. Tel: +886-2-23630231 ext. 4118; Fax: +886-2-23620879; E-mail: bocharng@ccms.ntu.edu.tw