Bot. Bull. Acad. Sin. (2004) 45: 301-306
Onguso et al. Effects of partial ringing and heating of trunk on shoot growth and fruit quality of peach trees
Effects of partial ringing and heating of trunk on shoot growth and fruit quality of peach trees
Justus M. ONGUSO*, Fusao MIZUTANI, and A.B.M. SHARIF HOSSAIN
The Experimental Farm, Faculty of Agriculture, Ehime University, Hojo 498, Hattanji, Ehime 799-2424, Japan
(Received March 24, 2004; Accepted June 4, 2004)
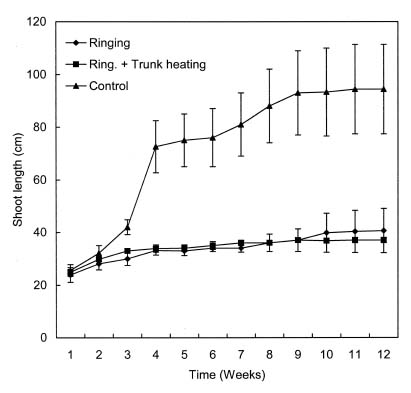
Abstract. Small-sized peach (Prunus persica [L.] Batsch.) trees for commercial fruit production have not been available due to the lack of suitable dwarfing rootstocks that have a wide range of compatibility among cultivars. Effects of partial girdling and trunk heating on the growth, yield, and fruit quality of peach were studied. A 4 cm wide partial ring of bark was removed at a height of 25 cm from the ground leaving a connecting strip of 5 mm. Furthermore, the de-barked spot was heated at 150°C for fifteen minutes using an electric heater. The partial girdling was effective in reducing shoot growth and improving fruit quality. The total soluble solids content of fruit in treatment trees was higher than that of control. The heating had little additive effects over partial ringing.
Keywords: Brix; Dwarfing techniques; Partial ringing; Starch content; Total sugar; Trunk heating.
Introduction
Small, compact, dwarfed, or size-controlled fruit trees seem to be the natural and obvious answer to many of the problems of commercial orchardists. They provide early fruiting, a more rapid turnover in varieties to meet changing market requirements, a lower cost of production, a higher proportion of high-grade fruit, and easier thinning, pruning, spraying, and harvesting (Tukey, 1978). The primary factor limiting the use of size-controlling rootstocks in stone fruit production is the lack of suitable rootstocks with a wide range of compatibility among cultivars (De Jong et al., 2001). This creates a need to explore alternative dwarfing techniques.
A number of workers have reported useful data on the application of various forms of girdling in fruit production. Ebell (1971) used overlapping, half _ circumference _ band girdles from which 25 mm wide strips of bark and phloem were removed from opposing sides of the stem of cone. Wheeler et al. (1985) compared partial _ overlapping _ band girdles to similar girdles applied with a pruning saw. They found both methods increased cone yield. Although stem girdling has received substantial attention, literature on the effects a combination of partial girdling and trunk heating has on shoot growth and fruit quality is limited. The interaction of bark width and shoot growth in peach has not been thoroughly investigated.
Partial ringing and trunk heating experiments were carried out to study their possible contribution to dwarfing in peach trees.
Materials and Methods
Site
The experiment was conducted at the Ehime University Experimental Farm located in southern Japan, 33°57' N, 132° 47' E at an elevation of about 20 m above sea level. The region has a mild temperate climate characterized by hot humid summers and cold dry winters. The soil at the experimental site is sandy loam (eutric fluvisol) with a pH (H20) of 5.7, a bulk density of a 1.08 g cm-3 and horizon A thickness of 0.15 m.
Plant Materials
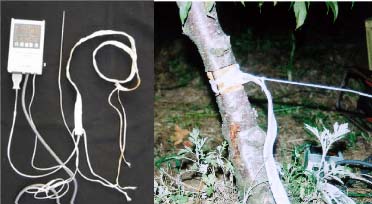
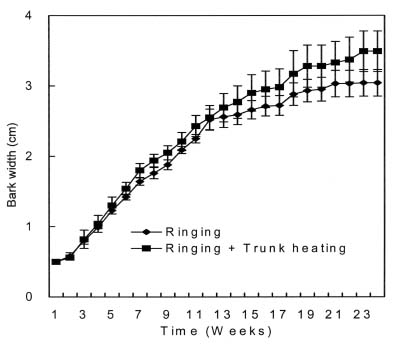
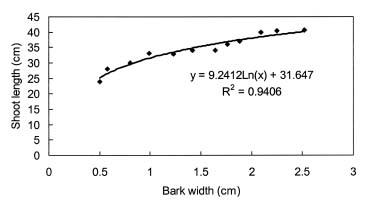
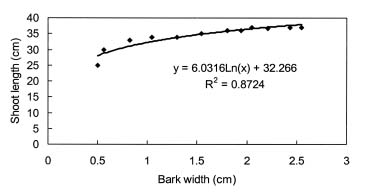
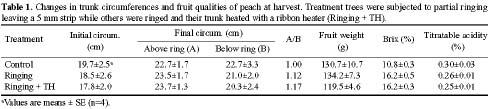
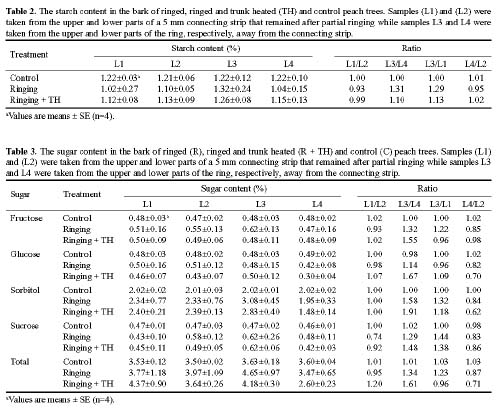
Four-year-old peach (Prunus persica Batsch. var. `Akatsuki x Banto') trees growing in an orchard at the University Farm were used in this study. On May 3, 2002, twelve trees were randomly selected and a 4 cm wide partial ring of bark removed from eight of them at a height of 25 cm above the ground to leave a 5 mm connecting strip. Four of the ringed trees were heated on the de-barked spot at 150°C for fifteen min using an electric ribbon heater (Figure 1). The growth of the trees was monitored weekly by measuring lengths of ten selected terminal shoots per tree. The growth of the 5 mm bark that was left after ringing was also monitored weekly using vernier calipers.
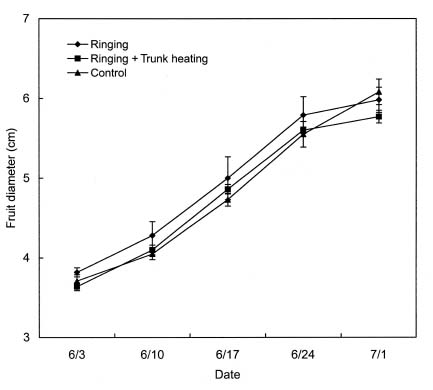
The fruit diameter was also evaluated weekly using vernier calipers. At harvesting, final fruit number, fruit diameter, and weight were recorded. Juice was extracted from the fruits and titratable acidity determined by acid-base titration using 0.1 N NaOH. The soluble solids content in the juice (Brix) was also measured by means of a refractometer (Atago PR-1).
*Corresponding author. E-mail: justusonguso@yahoo.com