Bot. Bull. Acad. Sin. (2004) 45: 307-313
Thomas and Puthur Regeneration in Kigelia pinnata L.
Thidiazuron induced high frequency shoot organogenesis in callus from Kigelia pinnata L.
T. Dennis THOMAS* and Jos T. PUTHUR
Postgraduate and Research Department of Botany, St. Thomas College, Pala, Arunapuram (P.O), Kottayam (Dt.), PIN- 686 574, Kerala, India
(Received December 24, 2003; Accepted June 29, 2004)
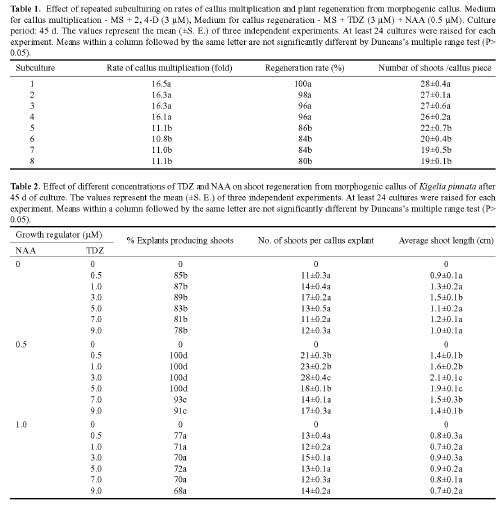
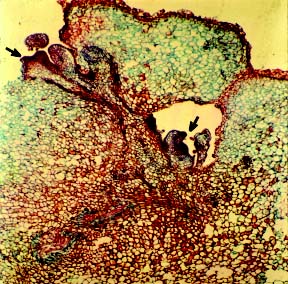
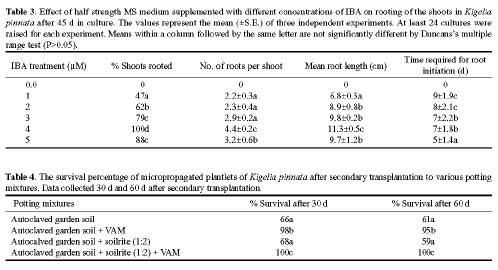
Abstract. An efficient regeneration system was developed for Kigelia pinnata L., a multipurpose tree belonging to the family Bignoniaceae. The nodal segments were cultured in vitro, and the optimum concentrations of plant growth regulators for callus induction were determined. The friable organogenic calli were derived from the basal cut end of the nodal segments. The highest yield of morphogenic callus (100%) was observed when nodal segments were cultured on Murashige and Skoog (MS) medium supplemented with 3 mM 2,4 dichlorophenoxyacetic acid (2, 4-D). The morphogenic callus maintained high regeneration during the first four subcultures in the callus induction medium. The maximum shoots (28/culture) were regenerated at the highest frequency of 100% when 3 mM thidiazuron (N-phenyl N' 1,2,3-thidiazol-5-yl urea) (TDZ) and 0.5 mM naphthaleneacetic acid (NAA) were added to MS medium. The emergence of multiple shoots from the calli was histologically documented. The regenerated shoots showed maximum rooting on ½ MS medium containing 4 mM indole-3- butyric acid (IBA). The effect of vesicular arbuscular mycorrhizae (VAM) association in averting the transplantation shock was tested and proved to be highly beneficial, giving a 100% survival rate after 60 d of transplantation. This efficient plant regeneration system provides a foundation for generating transgenic plants of this multipurpose tree.
Keywords: Kigelia pinnata L.; Medicinal plant; Plant regeneration; Thidiazuron; Tissue culture; Vesicular arbuscular mycorrhizae.
Abbreviations: 2, 4-D, 2,4 dichlorophenoxyacetic acid; IBA, Indole-3- butyric acid; NAA, Naphthaleneacetic acid; TDZ, thidiazuron (N-phenyl N' 1,2,3-thidiazol-5-yl urea); VAM, vesicular arbuscular mycorrhizae.
Introduction
Forest species predominate in temperate and equatorial zones, and the wood produced by the trees is the most abundant material on the earth's surface (Gammie, 1981). Wood provides fuel for most of the population of the world, particularly in developing countries, and is a leading industrial raw material. Despite its economic importance, the production of wood is threatened by population growth, desertification, industrial development, and attack by numerous parasites. The classical conservation techniques such as crossing, sexual and somatic hybridization, and breeding give a genetic blind mixture. These techniques are limited by the sterility of the descents, the genetic barrier between species, and the long life cycle of certain trees (Sederoff, 1995). Plant tissue culture offers many unconventional techniques for plant improvement. While tissue culture technology has been developed for the mass propagation of several fruit tree species, several other tree species are lagging behind due to a recalcitrant nature to in vitro techniques.
Kigelia pinnata, a fast growing, multipurpose tree used for ornamental and roadside planting, belongs to the fam
ily Bignoniaceae. Several parts of the plant are employed for medicinal purposes by certain aboriginal people. Traditional healers in India have used various parts of this plant to treat a wide range of skin ailments, from relatively mild complaints, such as fungal infections, boils, and psoriasis, to the more serious diseases like leprosy, syphilis, and skin cancer. Other medicinal applications include the treatment of dysentery, ringworm, tapeworm, post-partum haemorrhaging, malaria, diabetes, pneumonia, and toothache. An antimalarial compound known as lapachol has been extracted from the root of K. pinnata (Binutu et al., 1996). Another compound obtained from the wood, quinone, shows antimalarial activity against drug resistant strains of Plasmodium falciparum superior to chloroquine and quinine (Carvalho et al., 1988). Conventionally, K. pinnata reproduces via viable seeds, but the low percentage of seed viability limits its natural propagation. Hence alternative methods like in vitro techniques could be used to propagate this plant and thereby multiply elite genotypes. Micropropagation has many advantages over conventional propagation of fruit trees (Stushnoff and Fear, 1985) and is important for the regeneration following transformation (Ainsley et al., 2000) and cryopreservation (Channuntapipat et al., 2000). Thidiazuron (TDZ), a urea-derived cytokinin, is a potent cytokinin for woody plant tissue culture (Huetteman and
*Corresponding author. Telephone: +91-04822-212316,17; Fax: +91-04822-216313; E-mail: den_thuruthiyil@yahoo.com