Bot. Bull. Acad. Sin. (2004) 45: 315-324
Madhaiyan et al. Induction of systemic resistance by Methylobacterium
Growth promotion and induction of systemic resistance in rice cultivar Co-47 (Oryza sativa L.) by Methylobacterium spp.
Munusamy MADHAIYAN1,*, Selvaraj POONGUZHALI2, Murugaiyan SENTHILKUMAR2, Sundaram SESHADRI1, Heekyung CHUNG1, Jinchul YANG1, Subbiah SUNDARAM2, and Tongmin SA1
1Department of Agricultural Chemistry, Chungbuk National University, Cheongju, Chungbuk, 361-763, Republic of Korea
2Department of Agricultural Microbiology, Tamilnadu Agricultural University, Coimbatore 641 003, Tamilnadu, India
(Received December 9, 2003; Accepted June 8, 2004)
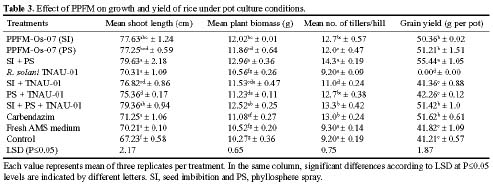
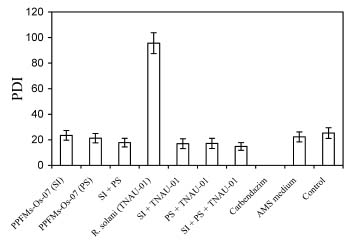
Abstract. Pink-pigmented facultatively methylotrophic bacteria (PPFMs), persistent colonizers of plant leaf surfaces, belong to the genus Methylobacterium and are mostly transmitted through seeds. Plant growth-promoting activity of methylotrophic bacteria and their effects on disease suppression were evaluated on rice under greenhouse conditions. Rice seeds were inoculated with Methylobacterium sp. strain PPFM-Os-07 and seed germination was evaluated in terms of morphometric measurements, seedling growth, rate of germination (RG), and seedling vigour index (SVI). Another experiment was carried out to study the induction of pathogenesis-related proteins (PR-proteins) in rice plants that were inoculated with methylotrophic bacteria by seed imbibition or foliar spray. In the third experiment, sixty-day-old rice plants grown in pots were challenge inoculated with Rhizoctonia solani strain TNAU-01. Methylobacterium inoculation promoted seed germination and plant growth. Increased plant height, number of tillers, plant biomass, and grain yield were observed. The average yield increases for seed imbibition and phyllosphere spray were, respectively, 22.1% and 24.3% greater than control. The bacteria also significantly reduced the sheath blight incidence when applied as either bacterial culture through seed imbibition and/or phyllosphere spray. The percent disease reduction recorded for seed imbibition alone and for combined applications of seed imbibition and phyllosphere spray were 17.8% and 23.5%. Rice plants sprayed with PPFM-Os-07 strain showed increased presence of PR-proteins and phenolic contents on day 1 after application. Maximum phenylalanine ammonia lyase (PAL) and peroxidase activity on day 4 and b-1,3-glucanase and chitinase activity on day 5 were recorded. The results suggest that Methylobacterium inoculation may alter rice susceptibility to R. solani. This work emphasizes the importance of evaluating induced systemic resistance while studying plant-associated growth promoting bacteria.
Keywords: Methylobacterium sp.; Oryza sativa L.; Induced systemic resistance; Pathogenesis-related proteins; Rhizoctonia solani.
Introduction
Methylobacterium spp. are a group of bacteria known as pink-pigmented facultative methylotrophs, or PPFMs (Austin and Goodfellow, 1979; Patt et al., 1976; Green and Bousfield, 1982, 1983). They are classified as alpha-Proteobacteria and are capable of growth on one-carbon compounds such as formate, formaldehyde, methanol, and methylamine as well as on a variety of C2, C3 and C4 compounds (Lidstrom, 2001). They can be easily isolated from plant tissues using selective media containing methanol as the sole carbon source (Corpe, 1985) and identified by their pink color, which distinguishes them from the other unrelated methylotrophic organisms normally encountered on plant tissue. They are abundant and non-pathogenic, distributed ubiquitously in the plant phyllosphere, and
have been isolated from more than 100 species of plants, ranging from liverworts and mosses to angiosperms and gymnosperms (Corpe and Basile, 1982; Dunleavy, 1990;1998 ). Hirano and Upper (1992) found them to represent greater than 90% of the phylloplane microflora of Phaseolus vulgaris throughout the growing season. PPFMs show populations ranging from 104 to 107 colony-forming units per gram fresh weight of plant tissue, with the highest numbers present on actively growing and meristematic tissue (Dunleavy, 1988; Corpe and Rheem, 1989; Holland and Polacco, 1992; Hirano and Upper, 1992; Holland, 1997). They are seed- transmitted (Corpe and Basile, 1982) and in dry soybean seeds they number 105 per gram (Dunleavy, 1988; Holland and Polacco, 1992). Their slow-growing nature and distribution over the whole plant suggest that their numbers are regulated simply by dilution as the plant tissue expands away from growing points. Holland and Polacco (1992) reported surface sterilization of seeds or other plant tissues, routinely carried
*Corresponding author. Fax: +82-43-271-5921; Phone: +82-43-261-2561; E-mail: mmadhaiyan@hotmail.com