Bot. Bull. Acad. Sin. (2004) 45: 325-330
Park et al. Molecular phylogeny of Monascus species
Phylogenetic relationships of Monascus species inferred from the ITS and the partial b-tubulin gene
Houng G. Park*, Elena K. Stamenova, and Shung-Chang Jong
American Type Culture Collection, P.O. Box 1549, Manassas VA 20108, USA
(Received February 16, 2004; Accepted June 14, 2004)
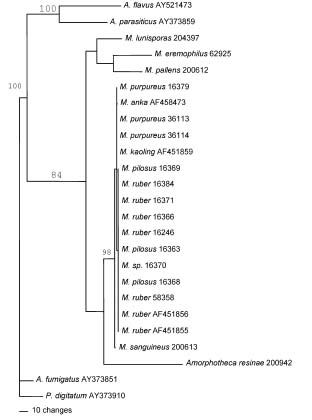
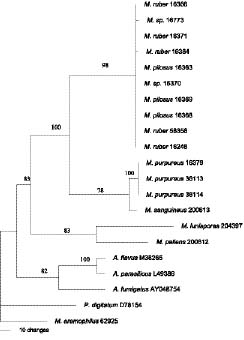
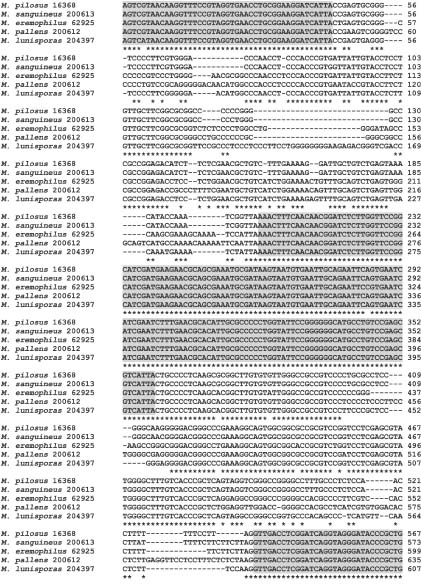
Abstract. ITS and partial b-tubulin genes of 17 ATCC reference strains of Monascus species were PCR amplified and sequenced. Monascus pilosus and M. ruber could not be differentiated with these sequences, suggesting a synonymy. In maximum parsimony analyses on both data sets, M. ruber, M. pilosus, M. purpureus, and M. sanguineus were placed into the same clade. ITS sequence alignment revealed a number of gaps in ITS1 and ITS2 of M. pallens, M. lunisporas, and M. eremophilus compared to M. purpureus, M. ruber, and M. pilosus. Accordingly, analyses with the ITS sequences placed these species into clades, incongruent with the analyses using the partial b-tubulin genes and the previous results with the partial large subunit rRNA genes. The phylogenetic relationship derived from the partial b-tubulin genes was similar to those postulated by the 5'-partial LSU rRNA genes. This finding strongly suggests that evolutionary or phylogenetic classification with ITS sequence information should be performed with caution. In the phylogenetic trees with the ITS sequences, M. lunisporas was distantly associated with Aspergillus ustus; M. pallens was placed in a clade that shares a common node with A. versicolor; and M. eremophilus was placed on a branch separate from the M. purpureus, M. ruber, and M. pilosus group while M. pallens and M. lunisporas were placed into the related clades sharing a common node in the tree derived from the partial b-tubulin gene. Each of the phylogenetic analyses with the partial b-tubulin genes, the ITS, or the 5'-end of the LSU rRNA, as previously carried out, placed M. eremophilus into a different lineage. Molecular analyses with these molecular targets generated three different topologies for M. eremophilus, indicating a unique and unpredictable genetic combination for this species. It might reflect extreme environmental stress on this species and subsequent genetic changes.
Keywords: b-tubulin; ITS; LSU rRNA; Monascus and phylogeny.
Introduction
Since the time Hawksworth and Pitt (1983) recognized three species of Monascus (M. pilosus K. Sato, M. ruber van Tieghem, and M. purpureus Went) based on physiological and morphological characteristics, several new species have been described. Barnard and Cannon (1987) described M. floridanus Cannon & Barnard, isolated from the roots of sand pine trees in Florida. Hocking and Pitt (1988) described a xerophilic species, M. eremophilus Hocking & Pitt, which differed from the other species by its slow growth rate, lack of an anamorph, and requirement for extremely dry conditions. Cannon et al. (1995) reported two additional species, M. pallens Cannon, Abdullah & Abbas and M. sanguineus Cannon, Abdullah & Abbas, based on the size of ascospores and colonies, pigmentation, and enzymatic activity tests using APIZYM strip tests (BioMerieux Vitek, Inc., Hazelwood, MO). In addition, Udagawa and Baba (1998) described M. lunisporas Udagawa & Baba, unique for its lunate ascospores and dark, olive-brown ascomata.
In 2003, phylogenetic relationships among the species were determined by sequences of the D1/D2 region of the large subunit (LSU) rRNA genes by Park and Jong. Monascus ruber and M. pilosus could not be differentiated. Monascus ruber, M. pilosus, and M. purpureus were closely related and clustered into the same subgroup.
We have carried out a further phylogenetic characterization using the ITS and partial b-tubulin genes in search of a better molecular differentiation marker and have evaluated the integrity and consistency of molecular phylogenetic relationships postulated by different molecular markers related to different biological functions.
Materials and Methods
Cultivation of the Strains
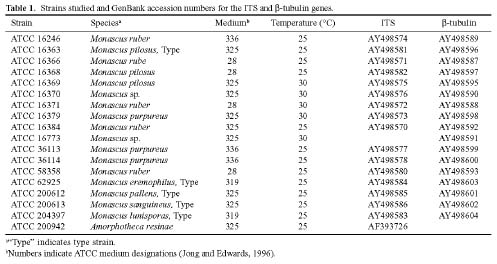
Seventeen strains of Monascus (Table 1) were obtained from cryopreserved material at ATCC. The strains were cultivated using one of four agar and broth media at 25°C or 30°C for approximately seven days. The media formulations included Blakeslee's formula (ATCC medium 325: malt extract 20 g, glucose 20 g, peptone 1 g, and agar 20 g per liter); PDA (ATCC medium 336: diced potatoes 300 g, glucose 20 g, and agar 15 g per liter); Emmon's modification of Sabouraud's agar (ATCC medium 28: Sabouraud's glu
*Corresponding author. Tel 703-365-2700 Ext 2562; Fax: 703-365-2730; E-mail: tohoungpark@yahoo.com