Bot. Bull. Acad. Sin. (2004) 45: 331-337
Chuang et al. Genetic diversity of non-heading Chinese cabbage in Taiwan
Genetic diversity and relationship of non-heading Chinese cabbage in Taiwan
Hsue-Yu Chuang1,3, Shing-Jy Tsao1, Jaw-Neng Lin2, Kan-Shu Chen2, Tsung-Dao Liou2, Mei-Chu Chung3, and Yau-Wen Yang3,*
1Department of Horticulture, National Taiwan University, Taiwan 106
2Fengshan Tropical Horticultural Experiment Station, Taiwan Agricultural Research Institute, Council of Agriculture, FengShan, Kaohsiung, Taiwan 830
3Institute of Botany, Academia Sinica, Nankang, Taipei, Taiwan 115
(Received February 16, 2004; Accepted June 8, 2004)
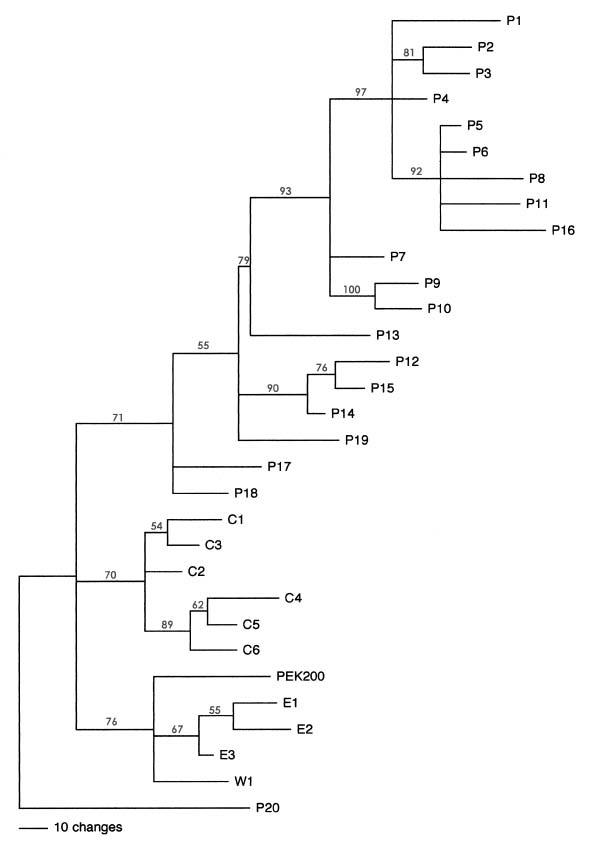
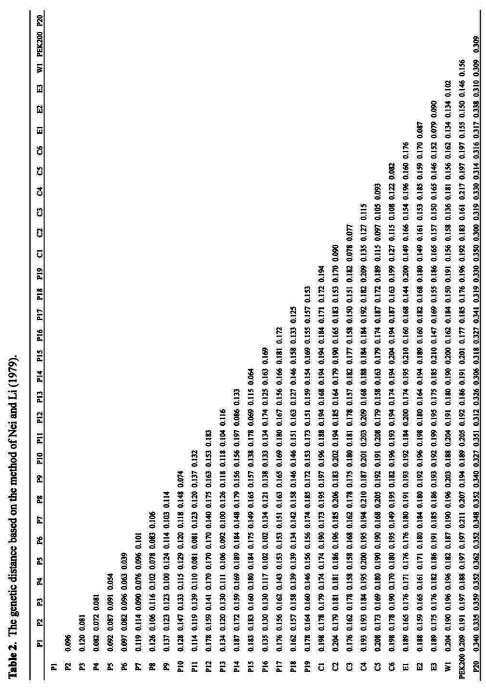
Abstract. DNA banding profiles generated by PCR reaction using different lengths of primers were used to study the genetic polymorphism and identities of 30 accessions from three major groups (pakchoi, chingansai, and edible rape) of Brassica rapa ssp. chinensis. Except for one accession later identified as B. juncea, all accessions can be separated into three major clades by applying maximum likelihood method as predicted by morphological taxonomy. Furthermore, the genetic relationships within different cultivar groups, and especially the pakchoi group, were also investigated based on their geography and growth habitat.
Keyword: Brassica rapa; Genetic diversity; Taiwan.
Introduction
Brassica rapa, one of the major economic Brassica crops, was probably introduced into China thousands of years ago and has since generated a rich morphological diversity in many subspecies and cultivar groups (Li, 1983; Opena et al., 1988). There are four major subspecies: chinensis, utilis, pekinensis, and rapifera (Ren et al., 1995). Of these, B. rapa ssp. chinensis (non-heading Chinese cabbage) is one of the most important vegetable crops in Taiwan. It accounts for 12% of total cultivated area for leaf vegetables (Taiwan Agriculture Yearly Report, 2001 in http: //www.coa.gov.tw/statistic/newyearbook/index.htm). The products of this subspecies translated into a market value of 2.7 billion NT dollars in 2001. Within chinensis, there are three major groups, pakchoi, chingansai, and edible rape, that account for many cultivars (including introductions) widely used in commercial year-round production in Taiwan. Although many cultivars were generated through intensive breeding in Taiwan, detailed documentation within this subspecies is often inadequate. In addition, some cultivars have lacked information about their genetic history since their introduction. Given the economic importance of these Brassica crops, improved methods are needed to appropriately identify the subspecies and group of each cultivar.
With the advent of recent methods in molecular biology, different molecular markers have been applied to the study
of phylogenetic relationships and identification among and within the Brassica species. These markers include restriction fragment length polymorphism (RFLP) (Song, 1988a, b; 1990; McGrath and Quiros, 1992), random amplified polymorphic DNA (RAPD) (Hu and Quiros, 1991; Quiros et al., 1991; Demeke et al., 1992; Kresovich et al., 1992; Mailer et al., 1994; Thormann et al., 1994; Ren et al., 1995; Lázaro and Aguinagalde, 1998; Divaret et al., 1999), and simple sequence repeat (SSR) (Kresovich et al., 1995; Charters et al., 1996; Szewc-McFadden et al., 1996; Westman and Kresovich, 1998; Plieske et al., 2001) among others. In this study, PCR analysis of DNA fingerprinting based on RAPD primers and simple sequence repeat (SSR) primers was used to generate molecular markers which were then used to study the genetic divergence and phylogenetic relationship among and within different cultivar groups in B. rapa ssp. chinensis. Based on these data, the corresponding relationships among geography, growing habit and known breeding history are discussed.
Materials and Methods
Plant Materials and DNA Extraction
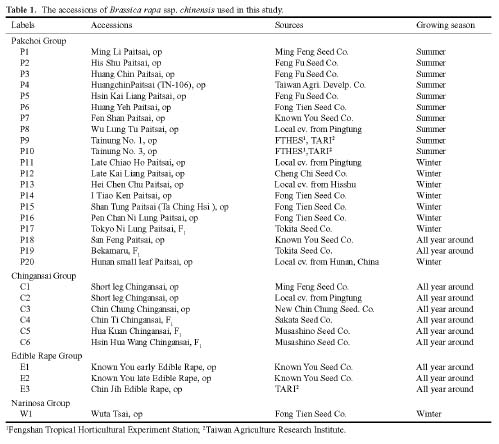
In Brassica rapa ssp. chinensis, twenty accessions of the pakchoi group, six accessions of the chingansai group, three accessions of edible rape, and one accession of the narinosa group, collected by the Fengshan Tropical Horticultural Experimental Station, were used in this study (Table 1). An accession of Chinese cabbage (PEK 200) from Chinglong Seed Co. was used as a comparison. Leaves
*Corresponding author. E-mail: yauwen@sinica.edu.tw