Bot. Bull. Acad. Sin. (2004) 45: 347-352
Chang and Chou Antrodia cinnamomea reconsidered and A. salmomea
Antrodia cinnamomea reconsidered and A. salmonea sp. nov. on Cunninghamia konishii in Taiwan
Tun-Tschu Chang1,* and Wen-Neng Chou2
1Division of Forest Protection, Taiwan Forestry Research Institute, 53 Nan-hai Road, Taipei 100, Taiwan
2National Museum of Natural Science, 1 Kuan-chien Road, Taichung 404, Taiwan
(Received February 17, 2004; Accepted May 24, 2004)
Abstract. Antrodia cinnamomea is reinstated as the correct name for a basidiomycete consistently associated with Cinnamomum kanehirai. Antrodia salmonea nov. sp., causing a brown heart rot of Cunninghamia konishii in Taiwan, is described and illustrated. Antrodia cinnamomea and A. salmonea are morphologically similar but can be separated by pore surface color of basidiomata and host preferences. Moreover, pairings between monokaryons of A. cinnamomea and those of A. salmonea demonstrate that these two fungi have different mating systems.
Keywords: Antrodia camphorata; Antrodia cinnamomea; Antrodia salmonea; Polypores; Taiwan.
Introduction
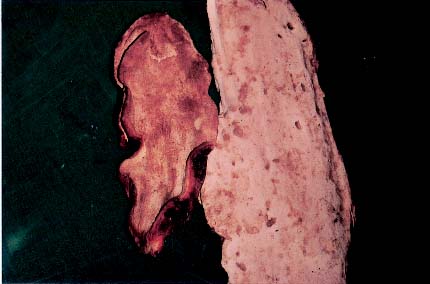
Cunninghamia konishii Hayata (Cunninghamieae) is a coniferous tree and endemic to Taiwan, growing in broad-leaved and coniferous forests at altitudes between 1,300 and 2,800 m in the central and northern parts of the island. A brown heart rot associated with resupinate, salmon-pink basidiomata in the empty rotten trunk of Cu. konishii has vernacularly been called Shiang-Shan-Chih. This fungus is similar to Antrodia cinnamomea T. T. Chang & W. N. Chou (Chang and Chou, 1995), which bears the vernacular name Niu-Chang-Chih and has only been collected from the endemic aromatic tree Cinnamomum kanehirai Hayata (Lauraceae) in Taiwan, but it has a different color on the pore surface of its basidiomata. The basidiomata of A. cinnamomea have medicinally been used for treatments of food and drug intoxications, diarrhea, abdominal pain, hypertension, skin itching, and cancer. Both species have a strong bitter taste, believed to indicate the presence of effective medicinal ingredients. Therefore, it is said that Shiang-Shan-Chih can substitute for Niu-Chang-Chih. In the present study, monokaryons (single basidiospore isolates) of the two species were paired to demonstrate that they have incompatible mating systems. A new name, Antrodia salmonea, is therefore given to the fungus associated with Cu. konishii.
Materials and Methods
Observation of Morphology
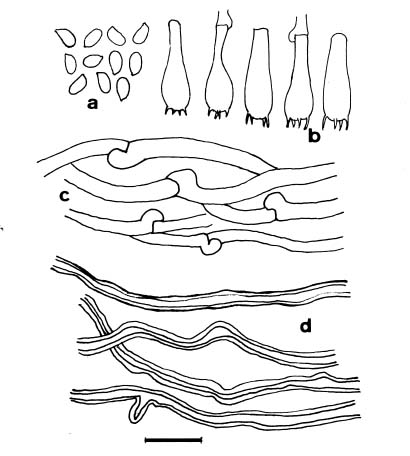
Descriptions of basidiomatal characters were based on fresh and dried specimens. Free-hand thin sections of basidiomata were mounted in two reagents for microscopic
studies: a 3% KOH solution was used to rehydrate the thin sections, and Melzer's reagent (IKI) was used to detect amyloidity and dextrinoidity.
Cultural Studies
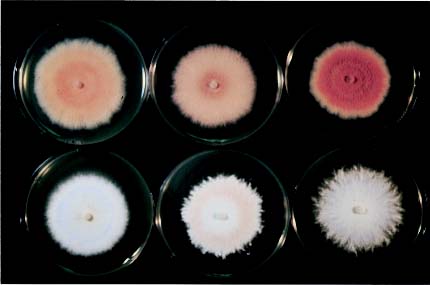
The methodology of Stalpers (1978) was followed for the cultural studies. The cultures were obtained from basidiomata and grown at 25°C on malt extract agar (MEA: 2% malt extract, 2% glucose, and 2% Bacto agar) and potato dextrose agar (PDA). The response to various temperatures was determined by measuring the linear growth of colonies on agar plate incubated at temperatures of 12, 16, 20, 24, 28, 32 and 36°C for 14 days. Petri dishes with an 85-mm internal diameter, which contained 20-25 ml agar media were inoculated with a piece of mycelium at the center, kept at 25°C, and macroscopically examined after 2 and 6 weeks of incubation. Separate slides prepared from various parts of the colonies were mounted in 3% KOH for observations by light microscopy.
Isolations of Monokaryons and Pairing Tests
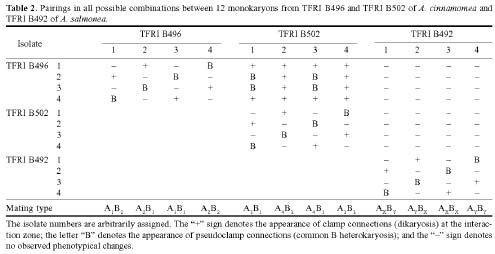
Basidiomata from two collections of A. cinnamomea (TFRI B496 from Tona, Kaohsiung and TFRI B502 from Alishan, Chiai) and from one collection of A. salmonea (TFRI B492 from Sheng-Kuaug, Taichung) were used for isolating monokaryons. Pieces of pore surface were cut from fresh basidiomata, placed into a test tube with sterile distilled water, and mechanically stirred for 5 min to obtain a basidiospore suspension. The basidiospore suspension was spread onto modified MEA (0.1% malt extract, 2% glucose, and 2% Bacto agar) plates, and monokaryons, i.e., colonies resulting from single basidiospores, were transferred to MEA after incubation at 24°C for 7-10 days.
Pairings among the monokaryons were made by placing small pieces of mycelia (2 mm in diameter) at a distance of 1 cm on MEA plates. Dikaryosis and common B
*Corresponding author. E-mail: ttchang@serv.tfri.gov.tw