Bot. Bull. Acad. Sin. (2005) 46: 1-10
HUNG et al. Hydrogen peroxide functions as a stress signal in plants
(Invited review paper)
Hydrogen peroxide functions as a stress signal in plants
Shu-Hsien HUNG1, Chih-Wen YU2, and Chin Ho LIN3,*
1Bioindustry Technology Department; 2Molecular Biotechnology Department, Da Yeh University, Taiwan
3Life Science Department, National Chung Hsing University, Taichung, Taiwan
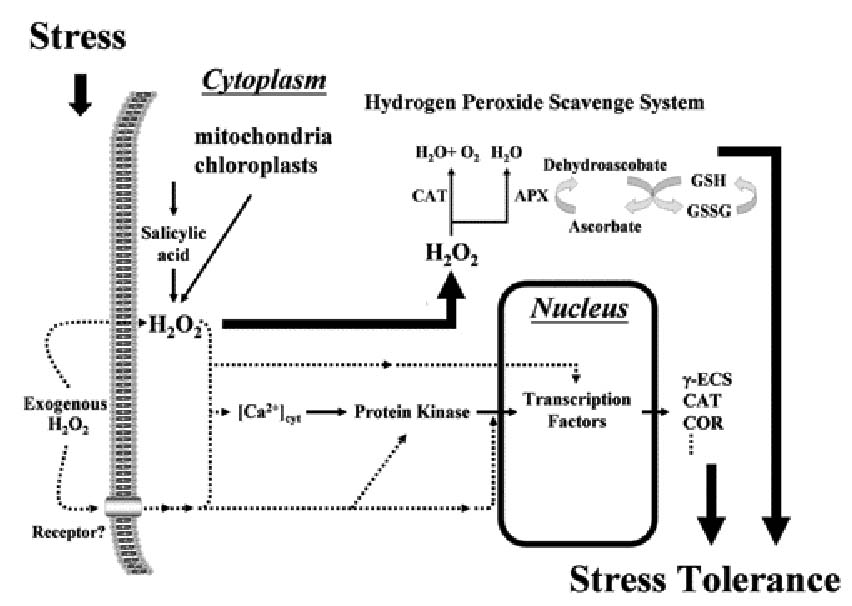
Abstract. Plants have evolved complex regulatory mechanisms in adapting to various environmental stresses. One of the consequences of many stresses is an increase in the cellular concentration of reactive oxygen species (ROS), which are subsequently converted to hydrogen peroxide (H2O2). An oxidative burst caused by biotic or abiotic stress leads to a disturbance in the cellular redox balance and is highly toxic to cells. Recently, H2O2, in addition to being a toxicant, has been regarded as a signaling molecule and a regulator of the expression of some genes in cells. These include genes encoding antioxidants, cell rescue/defense proteins, and signaling proteins such as kinase, phosphatase, and transcription factors. Here, we review the function of H2O2 as a signal molecule in the transduction of stress signals to the alteration of expression profiles of target genes, and we summarize the evidence that H2O2 acts as a stress signal in plants.
Keywords: Environmental stresses; Hydrogen peroxide (H2O2); Reactive oxygen species (ROS); Stress signal.
Abbreviations: ABA, abscisic acid; AP-1, activator protein-1; APX, ascorbate peroxidase; AsA, ascorbic acid; AtCBL1, Arabidopsis calcineurin-B-like protein; BSO, buthionine sulfoximine; CAT, catalase; CBF1, C-repeat binding factor; COR, cold regulated gene; CRT/DRE, C-repeat and dehydration-responsive element; DHAR, dehydroascorbate reductase; DREBs, DRE binding proteins; g-ECS, gamma-glutamylcysteine synthetase; GPX, glutathione peroxidase; GR, glutathione reductase; GSH, glutathione; GSSG, glutathione disulfide; GST, glutathione S-transferase; H2O2, hydrogen peroxide; HO·, hydroxyl radical; MAPKs, mitogen-activated protein kinases; MAPKKs, mitogen-activated protein kinase kinases; MAPKKKs, mitogen-activated protein kinase kinase kinases; MDHAR, monodehydroascorbate reductase; NF-kB, nuclear factor kB; NPK-1, Nicotiana protein kinase-1; O2·-, superoxide radical; PCD, programmed cell death; POX, peroxidase; PP2C, protein phosphatase 2C; PTPs, phosphotyrosine-specific phosphatases; ROS, reactive oxygen species; SOD, superoxide dismutase; UV, ultra-violet.
Contents
Introduction 1
The Generation and Removal of H2O2 2
H2O2 as a Signaling Molecule 3
Role of Ca2+ in H2O2 Signaling Pathway 4
The Role of Kinases and Phosphatases in the H2O2 Signaling Pathway 5
Transcriptional Regulation of Gene Expression in Response to H2O2 5
Conclusions 6
Literature Cited 6
Introduction
In nature, plants are exposed to various stresses, which affect their physiology, morphology, and development. Every year, environmental stress causes considerable losses in the productivity of many crops. Among these stresses, the fluctuation of temperature, the water status of soil, and the intensity of light are the most crucial signals affecting plant growth (Boyer, 1982; Trewavas and Malhó, 1997). In addition to external stimuli, a variety of
internal signals, such as hormones and nutrient conditions, also modify a plants metabolism, growth, and development. Hence, rapid and precise perception of and response to various stimuli are important as plants adapt to changing natural environments. However, how a plant perceives environmental changes and how it subsequently triggers signals to activate the physiological response are yet to be explored.
Extensive study on oxidative stress has demonstrated that exposure of plants to adverse environmental conditions induces the overproduction of ROS, such as superoxide radical (O2·-), H2O2, and hydroxyl radical (HO·) in plant cells (Wise and Naylor, 1987). ROS are highly reactive to
*Corresponding author. Tel: 886-4-22862511; Fax: 886-4-22862511; E-mail: chlin@mail.nchu.edu.tw