Bot. Bull. Acad. Sin. (2005) 46: 11-20
TUAN et al. Engineered baculovirus for better plant protection
Improved plant protective efficacy of a baculovirus using an early promoter to drive insect-specific neurotoxin expression
Shu-Jen TUAN1, 2, Roger F. HOU3, Suey-Sheng KAO1, Chi-Fen LEE1, and Yu-Chan CHAO2,*
1Biopesticide Department, Taiwan Agricultural Chemicals and Toxic Substances Research Institute, Wufeng, Taichung 413, Taiwan, ROC
2Institute of Molecular Biology, Academia Sinica, Nankang, Taipei 115, Taiwan, ROC
3Department of Entomology, National Chung Hsing University, Taichung 402, Taiwan, ROC
(Received April 5, 2004; Accepted September 30, 2004)
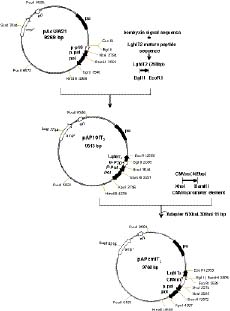
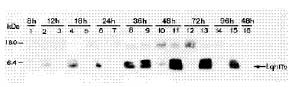
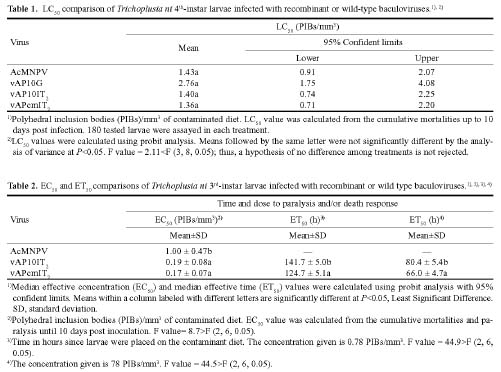
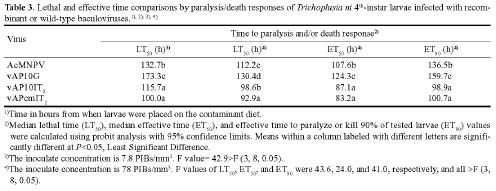
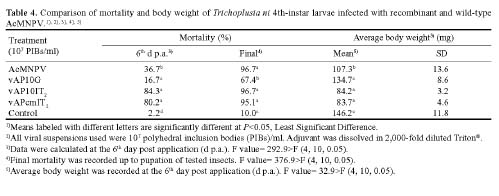
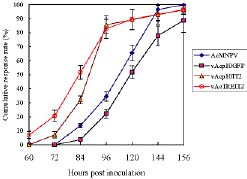
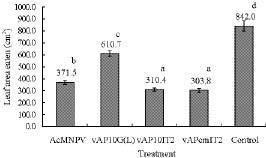
Abstract. Genetically improved Autographa californica (Speyer) multiple nucleopolyhedrovirus (AcMNPV), the type species of the genus Nucleopolyhedrovirus, was constructed with an insect-selective toxin gene (LqhIT2) derived from the venom of the Israeli yellow scorpion, Leiurus quinquestriatus (Ehrenberg). LqhIT2 was expressed by two distinct and temporally regulated promoters. These two promoters were the early promoter p-PCm (a pu-enhanced minimal CMV promoter) and the very late p10 promoter, resulting in recombinant viruses named vAPcmIT2 and vAP10IT2, respectively. Western blot and bioassay analysis showed that the toxin gene under the control of the early p-PCm promoter was expressed 24 h earlier than under the very late p10 promoter control, and vAPcmIT2 killed faster than vAP10IT2. The EC50 (effective concentration, EC) values of wild type AcMNPV, vAP10IT2, and vAPcmIT2 to the 3rd-instar larvae of Trichoplusia ni (Hübner) were 1.00, 0.19, and 0.17 polyhedral inclusion bodies (PIBs)/mm3, respectively. Compared to the very late promoter, the early promoter could shorten the effective time (ET50) by 14~17 h. Furthermore, for the 4th-instar larvae, compared to the wild-type virus and vAP10G, a 19~23% and 30~33% reduction in ET50 were found using vAP10IT2 and vAPcmIT2, respectively. Field trials of these two viruses showed an effective paralysis of the infected larvae which then stopped feeding, resulting in a decrease in the leaf area eaten compared to those larvae infected with wild-type virus and another control virus vAP10G, the GFP-expression recombinant AcMNPV. Scorpion toxin driven by the early p-PCm promoter shortened the time-to-paralysis of insect larvae, and thus provided an economical pest control advantage over that driven by the very late p10 promoter.
Keywords: AcMNPV; Autographa californica multicapsid nucleopolyhedrovirus; Baculovirus; Cabbage; Depressant neurotoxin; Leiurus quinquestriatus hebraeus; LqhIT2; Plant protection; Recombinant virus; Scorpion toxin; Trichoplusia ni.
Introduction
Since chemical pesticides frequently pose environmental and health risks, carefully selected biological control agents, like predators, parasitoids, and pathogens, have become attractive alternatives for the suppression of insect pests in the field. Baculoviruses (Baculoviridae) are arthropod-specific pathogens that have served as microbial biopesticides for control of many lepidopteran pests for several decades (Entwistle and Evans, 1985). These pathogens are characterized by large, circular double-stranded DNA genomes and rod-shaped virions that are enveloped within polyhedrin to form polyhedra (Tinsley and Kelly, 1985; Whitt and Manning, 1988). While baculoviruses have the potential for insect control, infected lepidopteran larvae still cause considerable crop damage due to continued feeding for several days after the initial infection, resulting in reduced efficacy in the field (Bonning and Hammock, 1996; Tuan et al., 1997; 1998).
A number of studies have established that the efficacy of baculovirus for insect pest control is greatly improved by the insertion of foreign genes into the viral genome. These include genes encoding the Manduca sexta (Linnaeus) diuretic hormone (Maeda, 1989), the Heliothis virescens (Fabricius) juvenile hormone esterase (Hammock et al., 1990), the Bacillus thuringiensis (Berliner) d-endotoxin (Martens et al., 1990; 1995), mite or spider toxins (Tomalski and Miller, 1991; Hughes et al., 1997; Priknod'ko et al., 1998), proteases (Harrison and Bonning, 2001), and the scorpion insect-specific neurotoxins (Stewart et al., 1991; Maeda et al., 1991; McCutchen et al., 1991; Fuxa et al., 1998; Gershburg et al., 1998; van Beek et al., 2003). All these studies reveal the significant contribution of exogenous gene products to the efficacy of baculovirus in pest control.
The LqhIT2 depressant insect toxin, derived from the Israeli yellow scorpion Leiurus quinquestriatus hebraeus (Ehrenberg), is a polypeptide of 61 residues (Zilberberg et al., 1991). Injection of LqhIT2 toxin into Sarcophaga falculata (Pandelle) blowfly larvae induced symptoms typical of the transient excitatory effect that preceeds the onset of prolonged flaccidity. The larvae are immobilized and
*Corresponding author. Phone: 886-02-2788-2697; E-mail: mbycchao@imb.sinica.edu.tw