Bot. Bull. Acad. Sin. (2005) 46: 21-28
HUNG and KAO Nitric oxide counteracts H2O2 action
Nitric oxide counteracts the senescence of rice leaves induced by hydrogen peroxide
Kuo Tung HUNG and Ching Huei KAO*
Department of Agronomy, National Taiwan University, Taipei, Taiwan, Republic of China
(Received February 17, 2004; Accepted July 26, 2004)
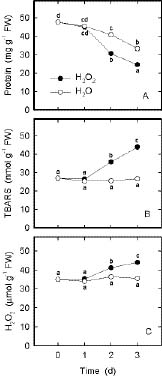
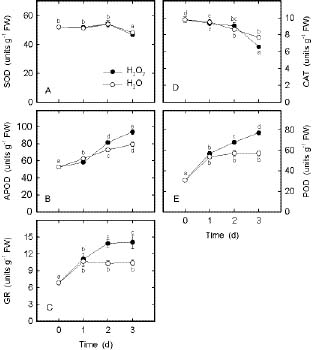
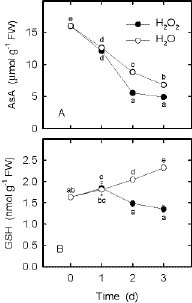
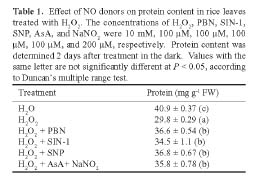
Abstract. In the present study, we evaluate the protective effect of nitric oxide (NO) against the senescence of rice leaves promoted by hydrogen peroxide (H2O2). Senescence of rice leaves was determined by decreases in protein content. H2O2 treatment resulted in (1) increases in leaf H2O2 content, (2) induction of leaf senescence, (3) increases in lipid peroxidation, (4) decreases in ascorbic acid (AsA) and reduced form glutathione (GSH) contents, and (5) increases in antioxidative enzyme activities (ascorbate peroxidase, glutathione reductase, and peroxidase). NO donors [N-tert-butyl-a-phenylnitrone (PBN), sodium nitroprusside, 3-morpholinosydonimine, and AsA + NaNO2] were effective in reducing H2O2-induced leaf senescence. PBN prevented H2O2-increased H2O2 content, H2O2-induced lipid peroxidation, H2O2-decreased AsA and GSH contents, and H2O2-increased antioxidative enzyme activities. The protective effect of PBN on H2O2-promoted senescence, H2O2-increased H2O2 content and lipid peroxidation, H2O2-decreased AsA and GSH contents, and H2O2-increased antioxidative enzyme activities was reversed by 2-(4-carboxy-2-phenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide, a NO-specific scavenger, suggesting that the protective effect of PBN is attributable to the NO released. Reduction of H2O2-induced senescence by NO in rice leaves is most likely mediated through its ability to scavenge H2O2.
Keywords: Hydrogen peroxide; Lipid peroxidation; Nitric oxide; Oryza sativa.
Introduction
Hydrogen peroxide (H2O2) is a constituent of oxidative plant metabolism and is itself an active oxygen species (AOS). H2O2 can also react with superoxide radicals to form more active hydroxyl radicals in the presence of trace amounts of Fe or Cu (Van Breusegem et al., 2001). The hydroxyl radicals initiate self-propagating reactions leading to peroxidation of membrane lipids and destruction of proteins (Halliwell and Gutteridge, 1989). H2O2 has been shown to promote leaf senescence (Parida et al., 1978; Mondal and Choudhuri, 1981; Sarkar and Choudhuri, 1981; Begam and Choudhuri, 1992; Lin and Kao, 1998), and induction of senescence is accompanied by an increase in endogenous H2O2 content (Mondal and Choudhuri, 1981). Lipid peroxidation is considered an important mechanism of leaf senescence (Thompson et al., 1987; Strother, 1988). The peroxidation of lipids can be initiated by active oxygen species (Thompson et al., 1987; Halliwell and Gutteridge, 1989). Thus, H2O2-induced leaf senescence is mediated, at least in part, through lipid peroxidation.
In mammalian systems, nitric oxide (NO), a bioactive molecule, is produced mainly from L-arginine by NO synthase (NOS). However, in plants neither the protein nor the appropriate gene for NOS activity have been detected (Lamattina et al., 2003). Recent work demonstrates that
the mammalian NOS antibodies recognize many NOS-unrelated plant proteins (Butt et al., 2003), suggesting that inferring the presence of plant NOS using the immunological technique may be inappropriate. These results led to speculation that the plant NOS-like enzyme could be structurally different from the mammalian NOS. Recently, a pathogen-inducible plant NOS has been identified as a variant P protein of the mitochondrial glycine decarboxylase complex (Chandok et al., 2003; Wendehenne et al., 2003). Additionally, Guo et al. (2003) reported the presence of an NOS gene (AtNOS1) in Arabidopsis. AtNOS1 turned out to be a protein very similar to a group of bacterial proteins with putative GTP-binding or GTPase domains. Neither the variant P protein nor the purified AtNOS1 protein had sequence similarities to any mammalian NOS. It is increasingly evident that plant nitrate reductase also catalyzes a NAD(P)H-dependent reduction of nitrite to NO (Yamasaki et al., 1999; Rockel et al., 2002, Sakihama et al., 2002; Lamattina et al., 2003). Evidence is mounting that NO acts as an important messenger in plant physiological processes, including growth, development, and defense responses (Noritake et al., 1996; Gouvea et al., 1997; Dangl, 1998; Beligni and Lamattina, 2000, 2001; Neill et al., 2002; Pagnussat et al., 2002; Jih et al., 2003; Zhao et al., 2004). NO has been shown capable of counteracting the toxicity of paraquat and diquat, which are known to generate superoxide radicals, in potato leaves (Beligni and Lamattina, 1999a,b; 2002) and block H2O2 production induced by jasmonic acid in tomato leaves (Orozco-Cárdenas and Ryan, 2002). NO seems to be a potent antioxidant, and its
*Corresponding author. Fax: +886-2-23620879; E-mail: kaoch@ntu.edu.tw