Bot. Bull. Acad. Sin. (2005) 46: 29-34
Wang et al. Cryopreservation of in vitro-grown shoot tips of papaya
Cryopreservation of in vitro-grown shoot tips of papaya (Carica papaya L.) by vitrification
Yuan-Long WANG1, Ming-Jen FAN2, and Song-Iuan LIAW1,*
1Department of Life Sciences, National Chung-Hsing University, 250, Kuo-Kuang Road, Taichung, Taiwan 40227, ROC
2Senior Scientist, National Plant Genetic Resources Center, Taichung, Taiwan, ROC
(Received March 12, 2004; Accepted July 14, 2004)
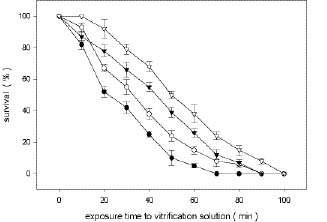
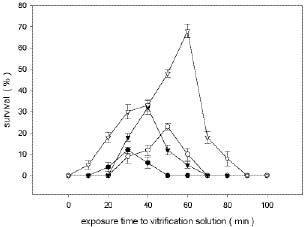
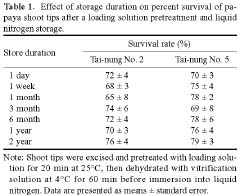
Abstract. Shoot tips from 4- to 6-week-old in vitro grown plantlets of six different cultivars of papaya were cryopreserved by vitrification. Shoot tips were treated in a 2 ml cryo-tube filled with a solution of 2 M glycerol and 0.4 M sucrose at 25°C for 20 min, then dehydrated with 1 ml pre-cooled vitrification solution at 4°C for 60 min. Treated samples were kept in liquid nitrogen for at least one week. The recovery percentages of six cultivars' shoot tips after vitrification were between 48% and 93%. Cryopreserved shoot tips after being warmed resumed growth within 7 days and developed shoots directly without intermediate callus formation. Prolonged storage in the liquid nitrogen up to 2 years had no affect on regeneration. This study has successfully developed a simple and effective protocol for the cryopreservation of papaya shoot tips.
Keywords: Carica papaya; Cryopreservation; Papaya; Vitrification.
Abbreviations: BA, 6-Benzylaminopurine; DMSO, Dimethylsufoxide; IBA, Indole-3-butyric acid; LS, Loading solution.
Introduction
Promoting sustainable utilization of biological resources is critical to the development of modern biotechnology. Because plant germplasm is indispensable for breeding new cultivars that enable the maintenance of genetic diversity and ecological environment and the avoidance of genetic erosion (Lambardi et al., 2000), germplasm conservation plays an important role in the sustainable utilization of biological resources.
Traditional methods of preserving plant germplasm are seed storage and maintaining plants in the field (Thinh et al., 1999). Those ways apply to most cultivars but have many disadvantages. Field collections require a large space and considerable labor, and the quantity of the species that can be preserved is limited (Hirai and Sakai, 1999; Lambardi et al., 2000; Pennycooke and Towill, 2000; Matsumoto et al., 2001). Field collections are also subject to environmental stress, climate change, and various plant diseases (Hirai and Sakai, 1999; Pennycooke and Towill, 2000; Matsumoto et al., 2001).
In vitro culture techniques supplement plant conservation and have been applied to germplasm collection, preservation, and rapid clonal multiplication. Due to factors such as the methods and duration of storage and the characteristics of the species, in vitro culture systems provide only short-medium term storage. Short-medium term
storage achieves conservation by delaying growth and increasing the time interval between culture generations, but this is laborious and time consuming (Engelmann, 1991; Escobar et al., 1997; Charoensub et al., 1999). In addition, genotypes could possibly be lost due to contamination or human error during the process (Engelmann, 1991; Thinh et al., 1999; Vandenbussche et al., 2000). Therefore, in vitro culture cannot be efficiently applied to long-term plant germplasm conservation.
For long-term conservation of plant germplasm, cryopreservation is currently the only option (Engelmann, 1991; Takagi et al., 1997; Thinh et al., 1999; Tsukazaki et al., 2000; Sakai, 2000; Matsumoto et al., 2001; Helliot et al., 2002). Storing plant seed, organs, tissues and cells in liquid nitrogen (-196°C) halts metabolic activities yet maintains their viability. In theory, this method enables plant materials to be stored indefinitely without physiological alteration or generation of somaclonal variation, and it requires minimal storage space and maintenance (Charoensub et al., 1999; Hirai and Sakai, 1999; Thinh et al., 1999; Sakai, 2000; Matsumoto et al., 2001; Touchell et al., 2002).
Traditional cryopreservation often uses a slow cooling to avoid intracellular ice formation, a common cause of lethal cell damage (Thinh et al., 1999; Lambardi et al., 2000; Touchell et al., 2002). However, equipment is costly, and the method is not effective for low temperature sensitive species (Pennycooke and Towill, 2000). Recently, approaches to plant germplasm cryopreservation involving direct plunging into liquid nitrogen have been explored,
*Corresponding author. Tel: 886-04-22840417-516; Fax: 886-04-22874740; E-mail: siliaw@mail.nchu.edu.tw