Bot. Bull. Acad. Sin. (2005) 46: 61-69
QIU et al. Genetic variation in Dysosma versipellis
A preliminary study of genetic variation in the endangered, Chinese endemic species Dysosma versipellis (Berberidaceae)
Ying-Xiong QIU1, Xin-Wen ZHOU1, Cheng-Xin FU1,*, and Yuk-Sing Gilbert CHAN2
1Laboratory of Systematic and Evolutionary Botany, Department of Biology, College of Life Sciences, Zhejiang University, Hangzhou 310029, P. R. China
2Department of Applied Biology and Chemical Technology, The Hong Kong Polytechnic University, Hung Hom, Hong Kong
(Received December 23, 2003; Accepted September 30, 2004)
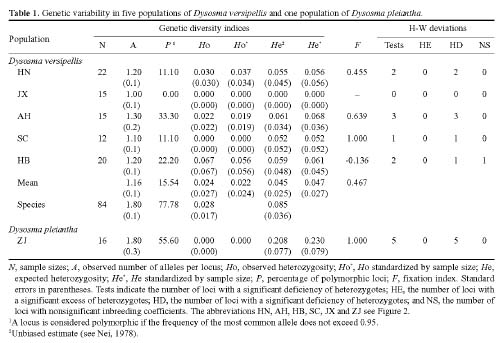
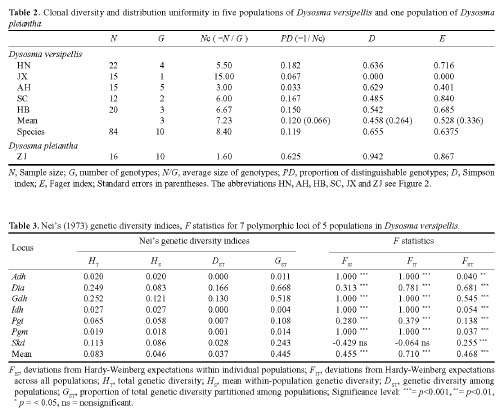
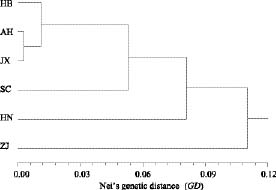
Abstract. This study represents a preliminary analysis of allozyme variation in Dysosma versipellis (Berberidaceae), an endangered plant species endemic to China. Five populations of D. versipellis and one population of D. pleiantha were sampled and analyzed using starch gel electrophoresis of nine enzymes that corresponded to nine interpretable loci. Levels of genetic polymorphism within populations (means: P = 15.54%, A = 1.16, He = 0.045) were much smaller than values for seed plants in general (P = 34.2%, A = 1.53, He = 0.113), as well as values for other endemic species (P = 26.3%, A = 1.39, He = 0.063). Mean values for the FST across all D. versipellis populations tended to be high (FST = 0.468). An indirect estimate of the number of migrants per generation (Nm = 0.284) indicated that gene flow is low among populations of D. versipellis. Additionally, analysis of genetic variation revealed a substantial heterozygosity deficiency in all analyzed populations except HB. Genotype frequencies within D. versipellis populations indicate that they may be severely inbred, making inbreeding depression a possible explanation for the low seed set observed in this species. Likewise, the low level of genetic diversity observed within D. versipellis populations suggests that clonal reproduction might be more important than sexual reproduction for D. versipellis. In comparison, genetic variation observed in one population of the closely related species D. pleiantha was much higher than the variation within D. versipellis populations. On the basis of these observations, we suggest that in situ conservation will be an important and practical measure for maintaining this species. If ex situ conservation is pursued, sampling should cover all populations across the species' distribution so as to retain as much genetic diversity as possible.
Keywords: Allozyme; Asexual reproduction; Dsma versipellis; Endangered species; Genetic variation.
Introduction
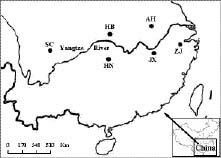
Plants in the genus Dysosma (Berberidaceae), with seven species, occur solely in China. Populations of the endangered species D. versipellis, an herbaceous perennial species that grows in the understory of mixed evergreen and deciduous forests (Ying et al., 1993), have a further restricted distribution in the East and South China. Dysosma pleiantha is another rare species in the genus, but little is known about its biology. However, it is clear that these two species have non-overlapping distributions and differ substantially in both leaf and flower morphology (Ying et al., 1993). These observations suggest that the breeding systems of some Dysosma species may have undergone divergent evolution resulting from adaptation to different ecological conditions. Dysosma versipellis reproduces sexually through cross-pollination and asexually by spreading rhizomes. Field observations indicates that although the plants flower almost every year, seed production is limited, and seedling establishment is poor.
Propagation, therefore, appears to occur mostly by vegetative means (Ma, 2000; Qiu and Qiu, 2002). In recent years, natural populations of this species have declined considerably due to anthropogenic activities like habitat destruction and overcollecting for medicinal applications. The rhizome of Dysosma has been found to be a source of podophyllotoxin, the active ingredient used as a starting compound for the chemical synthesis of etoposide (VP-16-213) and teniposide (VM-26), effective agents in the treatment of lung cancer, a variety of leukemias, and other solid tumor diseases (Jackson and Dewick, 1984, 1985). Nearly all remaining populations of D. versipellis are now located within protected nature reserves, and the species is classified as endangered in the Chinese Plant Red Book (Fu, 1992). Because the species is endangered, and harvesting of rhizomes continues to exceed the rate of natural regeneration, immediate attention should be given to conserving the species through in situ and/or ex situ approaches.
Resource managers responsible for the conservation and recovery of threatened plant species frequently need to make choices regarding which areas are in need of greatest protection, which populations should have priority for
*Corresponding author. Tel: +86-571-86971576; Fax: 86-571-86432273; E-mail: Biosys@zju.edu.cn