Bot. Bull. Acad. Sin. (2005) 46: 71-74
CHANG et al. Protocorm or rhizome?
Protocorm or rhizome? The morphology of seed germination in Cymbidium dayanum Reichb.
Chen CHANG*, Ying Chun CHEN, and Hsin Fu YEN
Department of Botany, National Museum of Natural Science, Taichung, Taiwan 404, Republic of China
(Received September 9, 2003; Accepted August 13, 2004)
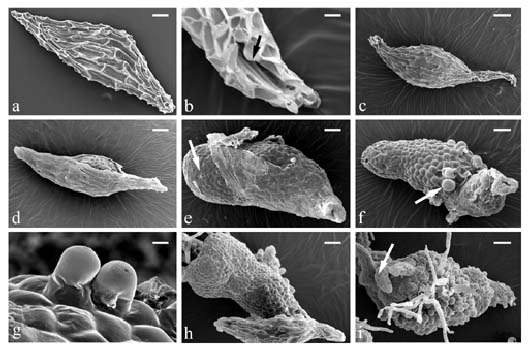
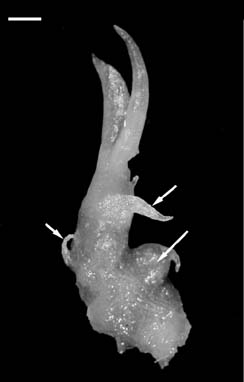
Abstract. Seeds of Cymbidium dayanum Reichb. were sown in vitro and germinated to produce protocorms one month later. Protocorms elongated, and the unicellular cell absorbing hairs were initiated at the base. The anterior of the protocorm produced the sheath leaf primordium. After two months, the protocorms grew, with the structure similar to that of rhizomes with nodes and lateral buds. The terminal buds spontaneously developed into shoots. According to the morphological pattern in vitro, we inferred that the seed germination of C. dayanum involved a fast transition from protocorm to plantlet by rhizome.
Keywords: Cymbidium dayanum Reichb.; Gravitropism; Protocorm; Rhizome.
Introduction
Under natural conditions, the seeds of all orchids germinate only after they are infected by their mycorrhizal fungi. In the laboratory, seeds of tropical epiphytic orchids germinate with relative ease on appropriate media. Seeds of terrestrial orchids from temperate regions are much more difficult to germinate in vitro (Arditti, 1992).
The structures that form between the germination of seeds and the establishment of seedlings have been denominated as protocorms or rhizomes. The term "protocorm" was first proposed by Melchior Treub in 1890 to describe the early stages in the germination of lycopods. Orchid protocorms resemble those of lycopodsaccording to Bermard's use of the termand it has been used extensively (Arditti, 1992). Protocorms have round or elliptical shapes with some unicellular absorbing hairs on the basal part and an apex meristem on the tip. The protocorms of most of tropical epiphytic orchids show the capacity to develop shoots directly. In many terrestrial orchids, the apices of protocorms elongate to form rhizomes that continue to grow and branch. The rhizome has several nodes, and the apex shows the physiology of gravitropism in vitro. After the elongation of the rhizome, the terminal bud grows upward and differentiates into shoots and roots.
Seedling establishment of the genus Cymbidium is distinguished by protocorm or by rhizome (Shimasaki and Uemoto, 1987). In epiphytic Cymbidium, protocorm directly develops into a shoot as, for example, C. dayanum Reichb., (Lu and Lee, 1990). Cymbidium aloifolium (L.) Sw. exhibits a fast transition from protocorm to plantlet by rhizome
(Nayak et al., 1998). In terrestrial Cymbidium, a rhizome needs 1-2 year to differentiate to a plantlet as, for example, C. ensifolium (L.) Sw. (Chung et al., 1985; Lu et al., 1992), C. forrestii Rolfe (Paek and Yeung, 1991), C. goeringii (Rchb.) Rchb. (Nagashima, 1982; Duan and Xie, 1983; Shimasaki and Uemoto, 1991), and C. sinense Willd. (Chiou and Wang, 1985; Chang and Chang, 2000).
This report describes the outer morphological changes during the seed germination of Cymbidium dayanum, an epiphytic Cymbidium, and indicates the morphology of intermediate structures between seed and plantlet.
Materials and Methods
Seed Germination
Capsules of Cymbidium dayanum were harvested 12 months after pollination and then sterilized with 2% NaOCl supplemented by one drop of Tween 20 in 15 min. The seeds were sown in 20×150 mm pyrex tubes each with 9 ml gelrite-medium that contained 1/4 MS salts (Murashige and Skoog, 1962), 0.5 mg l-1 niacin, 0.5 mg l-1 pyridoxine HCl, 0.1 mg l-1 thiamine HCl, 100 mg l-1 myo-inositol, 1 g l-1 peptone, 20 g l-1 sucrose, 2 g l-1 activated charcoal, 50 g l-1 banana pulp, and 4 g l-1 gelrite with a pH of 5.5. The cultures were exposed to artificial light of 1000 lux with a light/dark cycle of 16/8 h at 25°C±1°C.
SEM
Samples of seeds germination were fixed in 2.5% glutaradehyde in a 0.1 M phosphate buffer for 4 h, dehydrated through an ethanol series, dried in a critical-point dryer (Hitachi, HCP-2), and coated with gold in an ion coater (Hitachi, E1010). A Hitachi S-3000N scanning electron microscope was used to examine the samples.
*Corresponding author. Tel: 886-4-23226940 ext 153; Fax: 886-4-23285326; E-mail: cchang@mail.nmns.edu.tw