Bot. Bull. Acad. Sin. (2005) 46: 75-81
LIAO et al. Cuscuta pollen morphology in Taiwan
Pollen morphology of Cuscuta (Convolvulaceae) in Taiwan
Gwo-Ing LIAO1,*, Ming-Yih CHEN2, and Chang-Sheng KUOH1
1Department of Life Sciences, National Cheng-Kung University, Tainan 701, Taiwan
2Department of Life Sciences, National Chung-Hsing University, Taichung 402, Taiwan
(Received March 8, 2004; Accepted August 31, 2004)
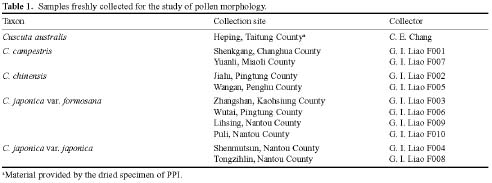
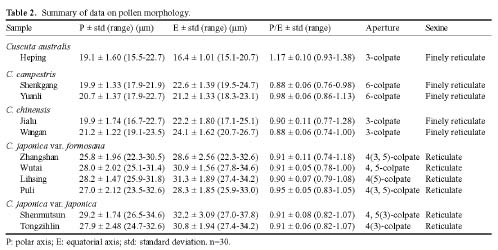
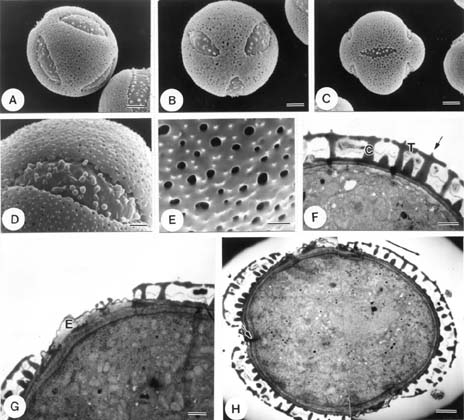
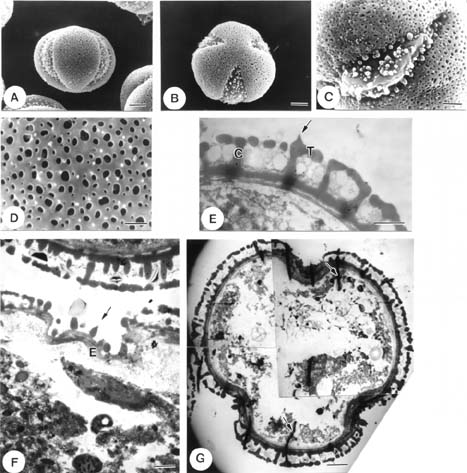
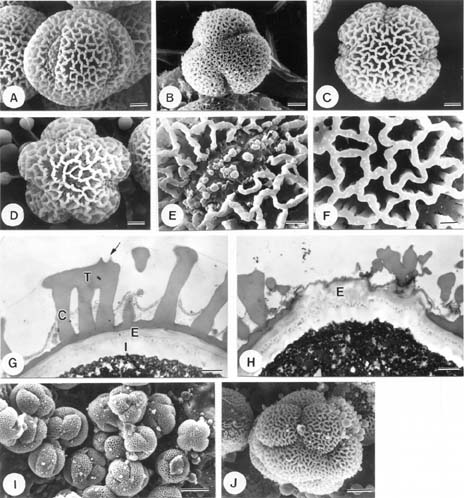
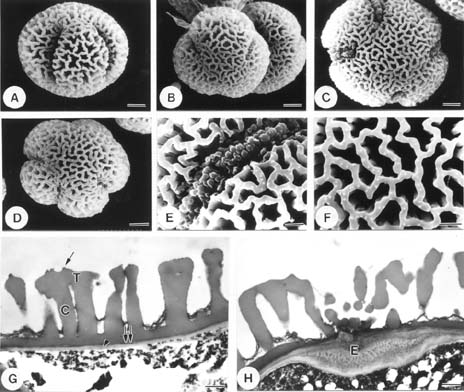
Abstract. The pollen morphology of five taxa of Cuscuta, C. australis, C. campestris, C. chinensis, C. japonica var. formosana and C. japonica var. japonica (Convolvulaceae) in Taiwan was investigated by using LM, SEM and TEM. The pollen of Cuscuta is three to six colpate, zonocolpate or pantocolpate colpus with granules, tectum with scabrate processes, ektexine reticulate or finely reticulate, and endexine thinner than ektexine. Two distinct pollen types of Cuscuta spp. in Taiwan are recognized. Type 1 is small, and has colpus with granules, and ektexine finely reticulate, including C. australis, C. campestris and C. chinensis. Type 2 is medium in size, and has colpus with granules, scabrate processes on surface of granule and ektexine reticulate, including C. japonica var. formosana and C. japonica var. japonica. Pollen types correspond with Yuncker's subdivision of the genus Cuscuta, type 1 for subgenus Grammica and type 2 for subgenus Monogyna. Meanwhile, pollen characteristics are also helpful in identifying species.
Keywords: Cuscuta; Dodder; Pollen; Taiwan.
Introduction
Dodders (genus Cuscuta L.) are parasitic flowering plants of the Convolvulaceae family (Yuncker, 1932; Kuijt, 1969; Parker and Riches, 1993; Liao et al., 2000). They are, however, sometimes assigned to the Cuscutaceae family (Hadac and Chrtek, 1970; Chrtek and Osbornova, 1991). This genus is globally distributed, with most species in the tropics and subtropics, and some in the temperate regions (Beliz, 1986). The number of species varies between 100 and 200 (Yuncker, 1932; Beliz, 1986; Parker and Riches, 1993; Fang et al., 1995; Staples and Yang 1998). Liao et al. (2000) considered three species and two varieties of Cuscuta in Taiwan, including C. australis, C. campestris, C. chinensis, C. japonica var. formosana and C. japonica var. japonica.
The identification of Cuscuta depends mostly on the detailed characteristics of the flowers, including the shapes of perianth segments, shapes of stigma, number of styles, infra-staminal scales attached to the filaments, as well as the dehiscence of the capsule. The characteristics of the scales and fruits are often difficult to determine, even in fresh materials. Furthermore, even these characteristics are difficult to observe in a dried specimen (Parker and Riches, 1993). Cuscuta campestris, a North American species recently introduced in several countries of the Old World, has often been misidentified (Parker and Riches, 1993). In Taiwan, it has been misidentified as C. australis or C. chinensis (Liao et al., 2000).
Information concerning pollen morphology can be used in the analysis of fossil pollen (Moore et al., 1991; Martin, 2001), airborne pollen (Nilsson et al., 1977; Lewis and Vinay, 1983), and honey pollen (Crompton and Wojtas, 1993). It is also used as taxonomical characteristics (Hsiao and Kuoh, 1995; Perveen and Qaiser, 1998). This study describes several pollen morphological characteristics of Cuscuta in Taiwan. The aim is to provide more information on the fine structural characteristics, and a Key is prepared for the identification of Taiwan taxa of Cuscuta.
Materials and Methods
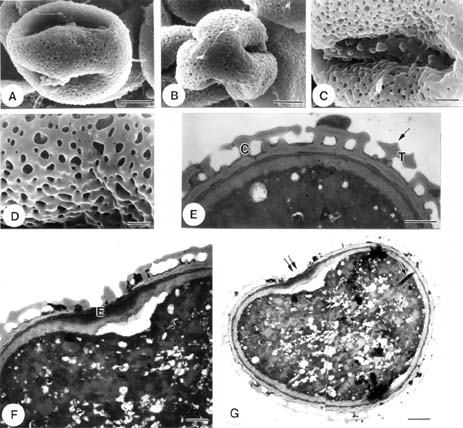
Eleven pollen samples representing five taxa of Cuscuta in Taiwan were examined by light microscopy (LM), transmission electron microscopy (TEM), and scanning electron microscopy (SEM). All pollen samples were freshly collected, except that of Cuscuta australia, which was obtained from the herbarium of National Pingtung University of Science and Technology, Taiwan (PPI) (Table 1).
Flowers were fixed using F. A. A. for 12-24 h and then stocked in 70% alcohol. For LM and TEM, anthers were dissected, gradually dehydrated in an acetone series, slowly infiltrated/embedded in Spurr epoxy resin, and then thick- and thin-sectioned on an ultramicrotome using a diamond knife. Thick sections (500 nm) were stained with toluidine blue, and examined on a Zeiss compound microscope. Ultrathin sections (90 nm) were stained with 4% aqueous uranyl acetate and 0.4% lead citrate, and examined/imaged with a Zeiss 10C TEM at 80 kV. For SEM, anthers were critical point dried. Pollen grains were then removed from the anthers, mounted onto aluminum stubs, coated with gold palladium, and then examined/imaged using a Hitachi H2500 SEM at 15 kV.
*Corresponding author. Tel: +886-6-2757575 ext 65540; Fax: +886-6-2742583; E-mail: liaomi@mail.ncku.edu.tw