Bot. Bull. Acad. Sin. (2005) 46: 107-118
KAO et al. AtMAPR, a novel protein for signaling
Characterization of a novel Arabidopsis protein family AtMAPR homologous to 25-Dx/IZAg/Hpr6.6 proteins
Ai-Ling KAO, Tsz-Yin CHANG, Shuo-Hsiu CHANG, Jong-Ching SU, and Chien-Chih YANG*
Department of Biochemical Science and Technology and Institute of Microbiology and Biochemistry, National Taiwan University, No. 1, Section 4, Roosevelt Road, Taipei, Taiwan, ROC
(Received July 28, 2004; Accepted October 11, 2004)
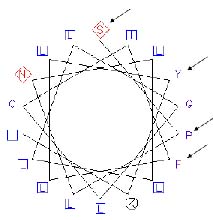
Abstract. Four Arabidopsis genes homologous to the membrane-associated progesterone binding protein (MAPR) were identified. MAPRs have previously been isolated from membrane preparations of porcine liver as proteins binding to progesterone. According to sequence alignment, each of the AtMAPRs, except AtMAPR2, was predicted to contain three domains; the N-terminal domain, except that of AtMAPR2, was predicted to accommodate a single transmembrane alpha-helix. A potentially interesting helical wheel motif SPX10FX2Y was found in the transmembrane domain. Proteins that may interact with AtMAPRs were found using the yeast two-hybrid system. AtMAPR2 and AtMAPR5DTM were found to associate with ubiquitin, where ubiquitination was involved in Aux/IAA modification. Sequence alignment indicated that AtMAPRs are distantly related to the Aux/IAA proteins. AtMAPR2 was also found to bind Myb3, a transcription factor that controls the expression of genes for the biosynthesis of phenylpropanoid. These results imply that AtMAPR may be a part of a plant hormone signaling pathway.
Keywords: AtMAPR; Non-genomic effect; Pant hormone signaling.
Abbreviations: ABA, abscisic acid; GA, gibberellic acid; MAPR, membrane-associated progesterone binding protein; AtMAPR, MAPR homologues in Arabidopsis.
Introduction
The components for signal transduction can be divided into two groups of proteins. These are, first, enzymes that are responsible for the biosynthesis of signaling molecules, or hormones, and second, proteins that are responsible for the perception of hormones, or receptors. The biosynthesis of major plant hormones has become ever clearer in the past 20 years. For example, most of the genes involved in the biosynthesis of gibberellins (GAs) are identified through genetic studies of dwarf cultivars or GA-deficient mutants (Hedden and Phillips, 2000). Many protein components for signal transduction were also determined through the genetic studies of GA-insensitive mutants, e.g. SPY, GAi and RGA (Sun, 2000), which were deficient in the GA signal transduction pathway. However, the receptors that can bind hormones directly, with the exception of the ethylene receptor and one receptor component for brassinosteroid, are still largely unknown. The perception of GA and ABA by membrane receptors has been proposed for nearly a decade, but these receptors still remain elusive (Lovegrove and Hooley, 2000). Data mining of a whole plant genome sequence to find membrane proteins as candidate receptors might offer an alternative approach to the genetic one.
Recently, the membrane receptor, or at least its major part, for brassinosteroids, BRI1, was identified and cloned by Chory's group from a series of BR-insensitive mutants (Wang et al., 2001). Brassinosteroids is similar in chemical structure to the steroid type hormones in animals. Thus, it seems feasible to compare the signaling pathways of steroids in animals and plants. Conventionally, steroid hormones including estrogens, progesterone, and androgens are thought to transmit signals through members of the nuclear receptor protein superfamily, namely estrogen receptor or progesterone receptor. This fact was mainly attributed to the hydrophobic nature of steroids. Steroid hormones diffuse into the cell and bind to the soluble nuclear receptors. In recent years, a different signaling pathway for steroids mediated by membrane-bound receptors has been demonstrated (Falkenstein et al., 2000). This type of signaling is not blocked by transcriptional inhibitors and is therefore described as the "non-genomic actions" of steroids.
A few proteins were proposed to be involved in the "non-genomic actions" of steroids, including MAPR, 25-Dx, IZAg, ratp28 and Hpr6.6, where sequence homology existed in these proteins (Falkenstein et al., 1996; Krebs et al., 2000; Nolte et al., 2000; Raza et al., 2001). Although its function was not fully understood, a membrane-associated progesterone binding protein (MAPR) was discovered from porcine liver cells (Falkenstein et al., 1996) and shown to be membrane-associated (Meyer et al., 1998). MAPR may undergo dimerization upon ligand binding
*Corresponding author. Tel: +886-2-23630231 ext. 3870; Fax: +886-2-23633783; E-mail: ccyang@ntu.edu.tw