Bot. Bull. Acad. Sin. (2005) 46: 119-125
LI et al. Protective effects of cobalt
The protective effects of cobalt on potato seedling leaves during osmotic stress
Chao-Zhou Li1, 3, Di Wang2, and Gen-Xuan Wang1,*
1College of Life Sciences, State Key Laboratory of Plant Physiology and Biochemistry, Zhejiang University, Hangzhou, Zhejiang Province 310029, P. R. China
2College of Agronomy, Gansu Agricultural University, Lanzhou 730070, P. R. China
3College of Life Sciences and Technology, Gansu Agricultural University, Lanzhou 730070, P. R. China
(Received July 6, 2004; Accepted November 18, 2004)
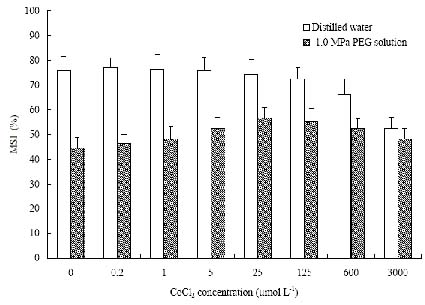
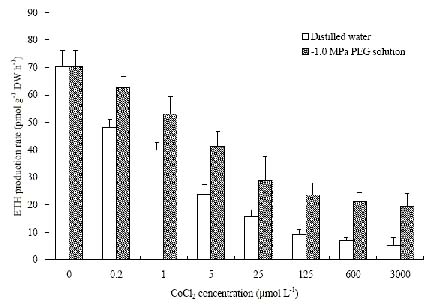
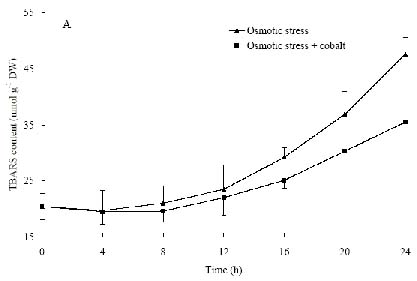
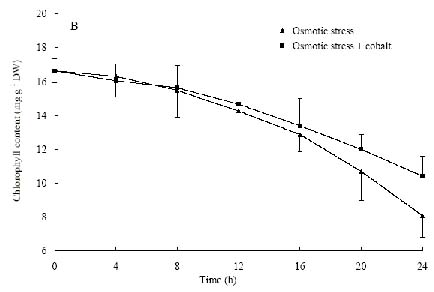
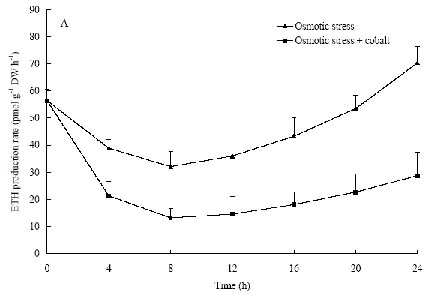
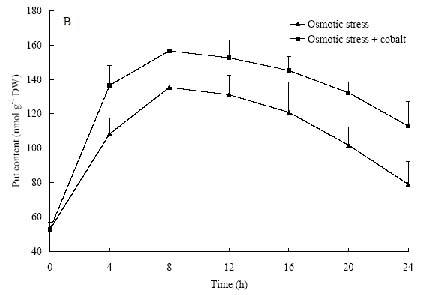
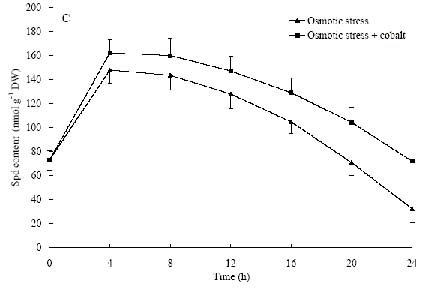
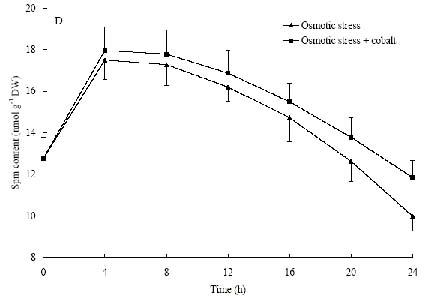
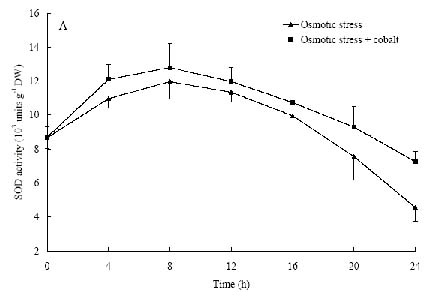
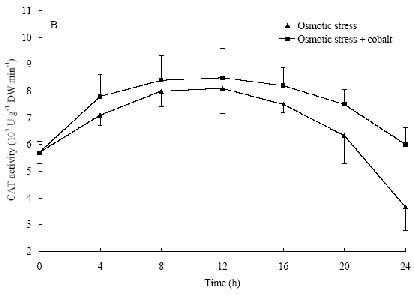
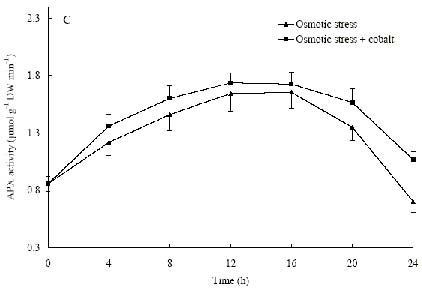
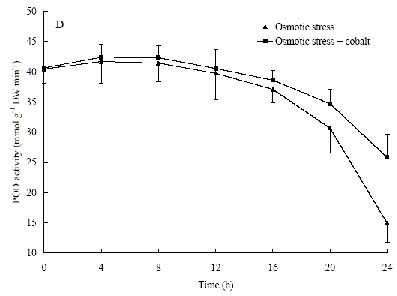
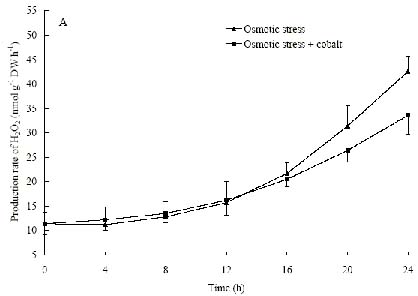
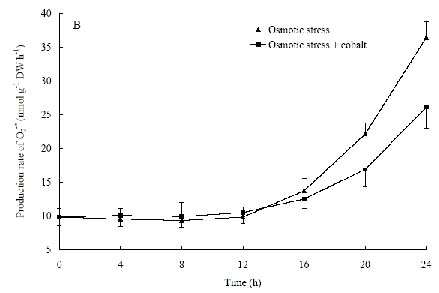
Abstract. The protective effects of cobalt on potato seedling leaves during osmotic stress were reported and discussed in this paper. The results showed that the membrane damage was significantly alleviated after 24 h of osmotic stress when cobalt of appropriate concentrations was added in PEG solutions. During -1.0 MPa PEG osmotic stress cobalt of 25 µmol L-1 inhibited ethylene production rate significantly. Cobalt of 25 µmol L-1 had no significant influence on the content of thiobarbituric acid reacting substances or chlorophyll in the first 16 h. When the stress lasted 20 h and 24 h, treatment with cobalt effectively restrained the increment in the levels of reactive oxygen species,hampered the decline in the content of putrescine, spermidine and spermine. The decline in the activities of anti-oxidative enzymes, thus inhibited the accumulation of thiobarbituric acid reacting substances and alleviated the reduction of chlorophyll content. It can be concluded that when the potato leaves were deeply stressed and damaged, through the inhibition of ethylene production, cobalt alleviated the decline in polyamine content and the activities of anti-oxidative enzymes, and hence alleviated the increment in reactive oxygen species levels and membrane damage and showed protective effects on the leaves.
Keywords: Cobalt; Ethylene; Osmotic stress; Polyamines; Potato.
Abbreviations: ACC, 1-aminocyclopropane 1-carboxylic acid; Co, cobalt; ETH, ethylene; MSI, membrane stability index; PAs, Polyamines; Put, putrescine; Spd, spermidine; Spm, spermine; ROS, reactive oxygen species; TBARS, thiobarbituric acid reacting substances.
Introduction
Cobalt (Co) is a kind of trace element and heavy metal found in soil (Hansen et al., 2001; Guevara et al., 2002) that can be incorporated into the active site of urease and render the enzyme inactive (Yamaguchi et al., 1999). Co concentration may be higher in serpentine, acidic, calcareous, or peaty soils, and may enter soils owing to the pollution of metal refineries and vehicular and aircraft exhausts (Freedman and Hutchinson, 1981). Excess Co induces yield reduction and an inhibition in assimilate production in leaves, and even inhibits the export of photoassimilates to roots and other sinks (Rauser and Samarakoon, 1980). Excess Co also causes oxidative stresses (Tewari et al., 2002) and may result in phytotoxity to plants (Chatterjee and Chatterjee, 2003). However, cobalt is unequivocally essential for leguminous crops as it is required for nitrogen fixation by bacteria in root nodules (Witte et al., 2002), and it even has beneficial effects on some non-leguminous crops (Locke et al., 2000).
Co is an inhibitor of 1-aminocyclopropane 1-carboxylic acid (ACC) oxidase and does inhibit ethylene (ETH) pro
duction (Lau and Yang, 1976; Locke et al., 2000). ETH shares the common precursor S-adenosylmethionine with spermidine (Spd) and spermine (Spm), so ETH and polyamines (Spd and Spm) are generally regarded to compete for a limited pool of S-adenosylmethionine (Wang et al., 2002). Furthermore, ETH may promote the oxidation of polyamines through its influence on reactive oxygen species (ROS) levels when osmotic stress became aggravated (Li et al., 2004). Since Co can reduce ETH production, we can hypothesize that it may have some influence on the content of polyamines, and even on membrane damage during environmental stress. However, to the best of our knowledge, little research has been conducted in this field, and the physiological effects of Co in proper concentration on the stressed plants remain unclear. Whether and how Co exerts its influence on cell membranes during environmental stresses needs to be investigated.
Materials and Methods
Plant Culture and Treatments
Potato (Solanum tuberosum L. cv. Gannongshu NO. 1) internodal segments were cultured in Murashige and Skoog (MS) medium which contained 30 g L-1 sucrose. The growth conditions were 22°C, 16-h photoperiod, and 120
*Corresponding author. Tel: +86-571-8697-1083; Fax: +86-571-8697-1083; E-mail: wanggx@zju.edu.cn (G.-X. Wang).