Bot. Bull. Acad. Sin. (2005) 46: 127-134
LI et al. SLA and LDMC in dune plants
Specific leaf area and leaf dry matter content of plants growing in sand dunes
Yulin LI1,*, Douglas A. JOHNSON2, Yongzhong SU1, Jianyuan CUI1, and Tonghui ZHANG1
1Cold and Arid Regions Environmental and Engineering Research Institute, Chinese Academy of Sciences, 260 Donggang West Road, Lanzhou, 730000, P.R. China
2USDA-ARS Forage and Range Research Laboratory, Utah State University, Logan, Utah 84322-6300, USA
(Received June 29, 2004; Accepted December 16, 2004)
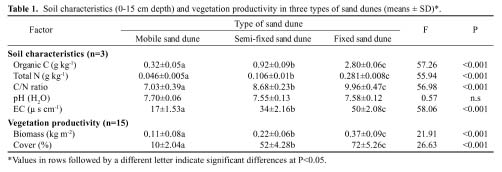
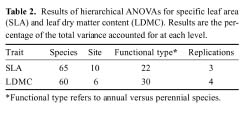
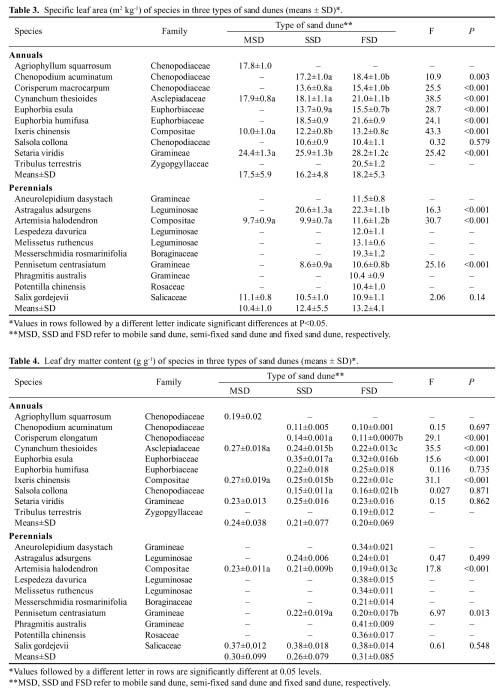
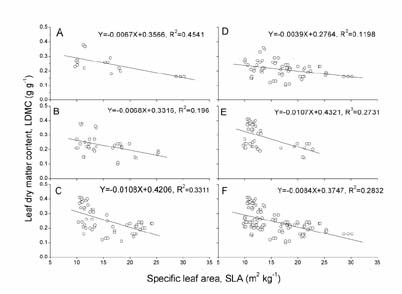
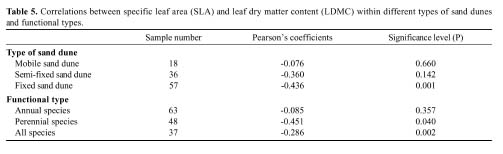
Abstract. We investigated the variations in specific leaf area (SLA) and leaf dry matter content (LDMC) of 20 species (10 annuals and 10 perennials) that have different distributional patterns in the Kerqin Sandy Land in northern China. The main purpose of our study was to determine if SLA and/or LDMC could be used as indicators of plant resource-use strategy in sand dune environments. The selected species were mainly distributed in three types of sand dunes: mobile sand dune, semi-fixed sand dune, and fixed sand dune. The three differed in their soil nutrient content and vegetation productivity. Both SLA and LDMC varied substantially among species in the different dunes. Maximum SLA (28.2 m2 kg-1) was observed in the annual grass Setaria viridis in a fixed sand dune, and the lowest SLA (8.6 m2 kg-1) was found in the perennial grass Pennisetum centrasiatum in a semi-fixed sand dune. LDMC varied from 0.10 g g-1 for the annual forb Chenopodium glaucum in a fixed sand dune to 0.41 g g-1 for the perennial grass Phragmitis australis in a fixed sand dune. For species common to two or three sites, their SLA and LDMC were significantly different between sites (P<0.05), except for two species for SLA and six species for LDMC. SLA and LDMC were negatively correlated both in the three types of sand dunes and for each functional type (annual versus perennial species). However, SLA and LDMC were significantly correlated only for the species in fixed sand dune, perennial species as well as all species. The results of our study showed that the SLA of annual plant species was generally higher than that of perennial species and that LDMC was significantly different between annual compared to perennial plants (P<0.05).
Keywords: Annuals; Leaf dry matter content; Perennials; Sand dune; Specific leaf area.
Introduction
Specific leaf area (SLA, leaf area per unit dry mass) and leaf dry matter content (LDMC, the ratio of leaf dry mass to fresh mass) are important traits in plant ecology because they are associated with many critical aspects of plant growth and survival (Garnier et al., 2001b; Shipley and Vu, 2002). Recent studies in controlled environments have demonstrated the important roles that these two traits play in explaining variation in potential relative growth rate (Poorter and Van der Werf, 1998) and ecological behavior in plants (Garnier et al., 2001a). According to a recent review (Poorter and de Jong, 1999), both SLA and LDMC are involved in the trade-off between rapid biomass production (high SLA, low LDMC species) and efficient conservation of nutrients (low SLA, high LDMC species). Because SLA and LDMC are indicator traits of resource-use strategies, it is important to evaluate these traits for different plant species in various environments (Westoby, 1998; Weiher et al., 1999).
Many studies have evaluated SLA and LDMC under both controlled and natural conditions involving various nutrient and water gradients (Cunningham et al., 1999;
Garnier et al., 2001a), vegetation types (Poorter and de Jong, 1999), plant functional types (Garnier et al., 1997), and leaf types (Vendramini et al., 2002). However, to our knowledge, few studies concerning these traits have been conducted on plants growing in sand dunes. Plants growing in sand dunes exhibit various morphological and ecophysiological characteristics (Liu et al., 1996). In the Kerqin Sandy Land of northern China, sand dunes can be classified into mobile sand dune, semi-fixed sand dune, and fixed sand dune according to the criteria proposed by Zhu et al. (1989). These sand dunes are widely distributed and alternate with gently undulating interdune lowlands to produce distinct differences in spatial heterogeneity of soil resources, which in turn leads to differences in species distribution, plant growth, and biomass production (Su and Zhao, 2003). These processes further amplify the differences among different types of sand dunes.
The main objective of the present study was to determine if SLA and LDMC differ in three types of sand dunes that occur in Kerqin Sandy Land of northern China, and evaluate whether SLA and/or LDMC could be used as indicators of plant resource-use strategy in these environments. In addition, we hoped to determine if annual and perennial plants differed in SLA and LDMC in the three types of sand dunes.
*Corresponding author. E-mail: liyleml@263.net