Bot. Bull. Acad. Sin. (2005) 46: 135-142
AHMAD et al. Salt tolerance of green gram genotypes at various growth stages
Comparative morphological and physiological responses of green gram genotypes to salinity applied at different growth stages
Shakil AHMAD1, Abdul WAHID2,*, Ejaz RASUL2, and Abdul WAHID1
1Department of Botany, Government College University Lahore, Pakistan
2Department of Botany, University of Agriculture, Faisalabad-38040, Pakistan
(Received August 27, 2004; Accepted December 20, 2004)
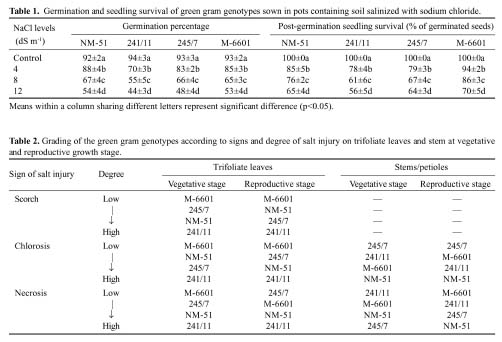
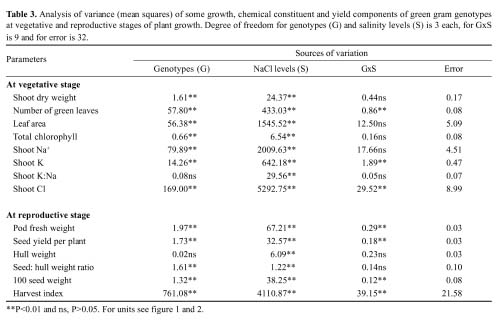
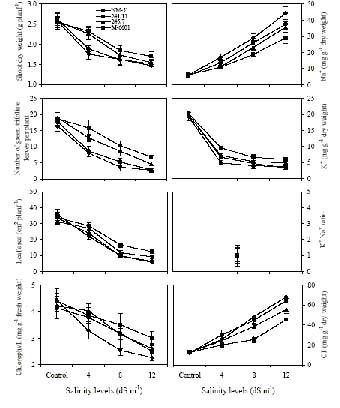
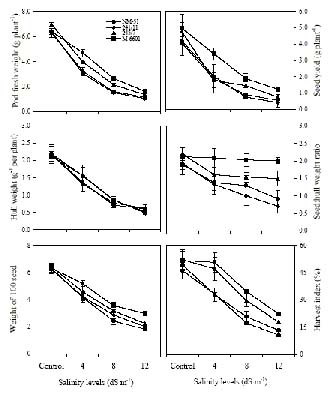
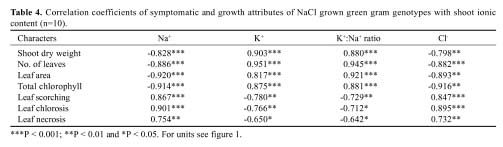
Abstract. Experiments were conducted to find the difference among green gram (Vigna radiata L. Wilczek) genotypes for salinity tolerance during germination and during vegetative and reproductive growth stages under NaCl treatment. Data revealed significant genotypic differences in germination percentage and post-germination survival of seedlings, symptoms of salt injury, changes in the levels of Na+, Cl- and K+ and total chlorophyll at the vegetative stage, and in pod development, seed yield, and yield components at the reproductive stage. Salinity tolerance in green gram was related to greater final germination and post-germination seedling survival, low scorching, chlorosis and necrosis of aerial parts, reduced Na+ and Cl-, slightly enhanced K+, and greater chlorophyll content. Appearance of increased symptoms on the aerial parts was positively related to increased Na+ and Cl- and negatively to increased K+. At maturity, pod weight, seed:hull weight ratio, 100-seed weight, seed yield and harvest index were also greater in M-6601. Maintenance of a steady seed:hull weight ratio in M-6601 indicated that a higher seed yield in this genotype is principally due to a greater partitioning of photoassimilates to seed rather than hull during pod development under saline conditions. Based on these criteria, M-6601 and 241/11 were declared salt tolerant and sensitive, respectively. Essentially then, ion-toxicity is the dominant factor modulating the salt tolerance of green gram during growth periods.
Keywords: Green gram; Harvest index; Ion-toxicity; Symptoms; Seed:hull weight ratio; Seedling survival.
Introduction
Rapidly increasing soil salinity has multifarious effects on plant growth and productivity. Salt-affected land comprises 19% of the 2.8 billion hectares of arable land on earth, and an increase in this menace is posing a serious threat to agriculture globally (Pessarakli and Szabolcs, 1999). Higher amounts of toxic ions in the root zone cause damage initially to roots and then to shoots after their transport. It is believed that greater ion injury at any stage of plant development is crucial for the maintenance of the active size of the canopy (Francois and Maas, 1999). Salt tolerance varies considerably with the developmental stages in a number of species (Wahid et al., 1997; Wilson et al., 2000). Tolerance at emergence followed by seedling survival and establishment are important in the maintenance of optimal crop stand in the field, and ultimately the economic yield (Wahid et al., 1999a; Raptan et al., 2001; Bayuelo-Jimenez et al., 2002).
The criteria used to appraise the salt tolerance potential of any plant species are morphological, physiological, and biochemical in nature (Rawson et al., 1988; Shannon, 1997; Flowers, 2004; Ashraf and Harris, 2004). The morphological criteria include stunted growth (Srivastava and Jana, 1984; Boyd and Rogers, 2004), leaf scorch (Barroso and Alvarez, 1997; Karakas et al., 2000), chlorosis of green parts (James, 1988; Pentalone et al., 1997; Husain et al., 2003),
and necrosis of leaves (Volkmar et al., 1998; Chen et al., 2003). Physiological criteria are tissue ionic contents and photosynthetic rate (Schachtman and Munns, 1992; Murillo-Amador et al., 2002; Morant-Manceau et al., 2004) while biochemical ones include qualitative and quantitative changes in proteins, fats, and carbohydrate patterns (Dubey, 1999; Khatkar and Kuhad, 2000; Bassil and Kaffka, 2002). Induced nutrient deficiency is one of the most important aspects of salinity, leading as it does to serious perturbation of normal cellular activities. The appearance of signs of salt damage is due to alterations in the nutrient status of tissues, which can be used to diagnose a stress response. For instance, deficiency of K+ leads to chlorosis followed by necrosis of leaves (James, 1988). Deficiency of N and Ca2+ leads to chlorosis (yellowing), and one of P results in necrotic spots on the leaf surface (Taiz and Zeiger, 2002). A majority of the criteria of salinity tolerance are physiological and biochemical (Ashraf and Harris, 2004), and only a few are concerned with the morphological changes at different stages (Wahid et al., 1999b; Husain et al., 2003; Boyd and Rogers, 2004). In addition, attributes like greater seed yield per plant and harvest index are adjudged as valid determinants of salinity response (Lawn and Rebetzke, 1991; Volkmar et al., 1998; Zeng et al., 2001; Bassil and Kuffka, 2002). Taken together, these factors can be used to predict the salinity response and to develop remedial strategies thereafter.
Green gram (Vigna radiata L. Wilczek) is an important traditional crop the world over. It is of short duration, re
*Corresponding author: E-mail: drawahid2001@yahoo.com