Bot. Bull. Acad. Sin. (2005) 46: 143-149
HSU et al. Anatomical characteristics of the secondary phloem in branches of Zelkova serrata
Anatomical characteristics of the secondary phloem in branches of Zelkova serrata Makino
Ying-Shan HSU1, Shiang-Jiuun CHEN1, Chin-Mei LEE2, and Ling-Long KUO-HUANG1,*
1Department of Life Science; Institute of Ecology and Evolutionary Biology, National Taiwan University, Taipei, Taiwan, Republic of China
2Division of Forest Utilization, Taiwan Forestry Research Institute, Taipei, Taiwan, Republic of China
(Received June 21, 2004; Accepted December 23, 2004)
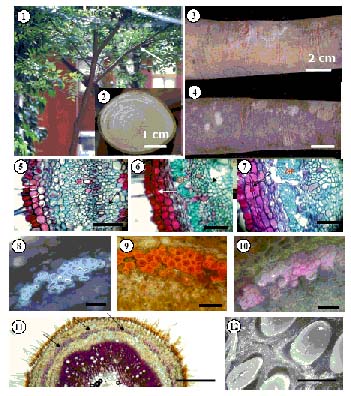
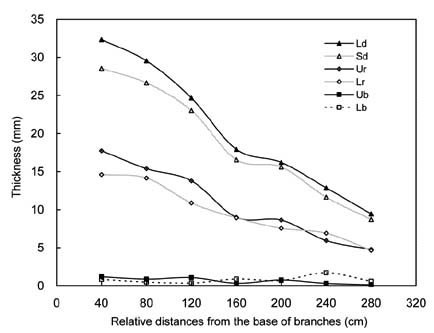
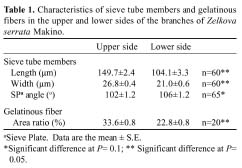
Abstract. The properties and tissue compositions of reaction wood in the leaning trunks or branches of trees have been extensively investigated, but studies on the influence of the related reaction on the secondary phloem are few and incomplete. In this work, formation of the vascular and cork cambium and the secondary phloem in the upper and lower sides of branches of Zelkova serrata Makino were studied. Vascular cambium is formed first in the second internode of shoot. In the third internode, cork cambium began to initiate subepidermally, meanwhile two to four layers of grouped gelatinous fibers were first found in the cortex outside of all the primary phloem. Gelatinous fibers were also found in both the secondary xylem and secondary phloem. The leaning branches exhibited pronounced radial secondary growth promotion to the upper side, and the reaction wood formed eccentrically. The secondary phloem formed layers of conducting sieve elements alternately with gelatinous fibers. Comparing the secondary phloem in the upper side (reaction phloem) of the branches with that in the lower side (opposite phloem), there is no obvious difference in thickness. Nevertheless, it was found that in the cross sections, the gelatinous fibers formed earlier, and there were more continuous cell seriates and a much larger area ratio in the upper side. Besides, the sieve tubes in the upper side of secondary phloem were longer and wider and possessed a very horizontally-orientated sieve plate between two sieve elements. These features may imply that, in branches of Zelkova serrata, the secondary phloem of the upper side (reaction phloem) may have a higher translocation efficiency.
Keywords: Branches; Gelatinous fibers; Reaction phloem; Secondary phloem; Zelkova serrata.
Introduction
With continued secondary growth, a tree's vascular cambium pertinently divides and produces secondary xylem inward and secondary phloem outward. The inclined trunk and branches frequently display eccentric growth and show specific structural changes in the cell layers, cell morphology, and cell wall constitution during the formation of secondary tissues (Fahn, 1990; Jourez et al., 2001). The timber which secondary growth slants toward one side is called reaction wood (Wilson and Archer, 1977). Generally, reaction wood is classified as compression wood or tension wood. Compression wood occurs in most gymnosperms, and tension wood forms mostly in the woody angiosperms (Wilson and Archer, 1977; Timell, 1986). Compression wood forms in the lower side of leaning trunks and branches and manifests its effect by slowly pushing the stem while tension wood forms in the upper side of leaning stems and functions by pulling the stems to the needed orientation. A tree, in effect, ingeniously adjusts its growth stress to the requirements of its environment, thereby satisfying its physical needs.
It is reasonable to expect the inductive causes of reaction wood formation to also affect the derivatives of the vascular cambium on the correlative phloem side. Scurfield and Wardrop (1962) first coined the term reaction phloem. The terms reaction, compression, and tension barks were coined by Timell (1986). The different morphology and structure of barks on the lower and the upper sides of branches or on the leaning stems of conifers have been mentioned by several investigators. In Chamaecyparis obtuse and Cryptomeria japonica, Onaka (1949) found radial growth in the bark to be larger on the lower, compression wood side than on the upper side although the difference was less than in the wood. As a consequence of the larger radial growth of the compression wood, outer bark on the lower side tended to split and fall off, and compression bark as a result was thin. Kutscha et al. (1975) observed that in compression phloem of Abies balsamea the bands of tannin cells were discontinuous instead of continuous as in normal phloem. Thick, unlignified wall layers have been observed in gelatinous fibers present in tension phloem, for example in Eucalyptus (Dadswell and Wardrop, 1955), Tilia cordata (Böhlmann, 1971), and Populus euramericana (Nanko et al., 1982).
Many studies have explored the structural, physiological, and mechanical properties of reaction wood (Wilson and Archer, 1977; Timell, 1986). However, the for
*Corresponding author. Tel: (02) 33662510; Fax: (02) 23673374; E-mail: linglong@ntu.edu.tw