Bot. Bull. Acad. Sin. (2005) 46: 151-154
Chang and Wang Basidiomatal formation of Antrodia cinnamomea on media
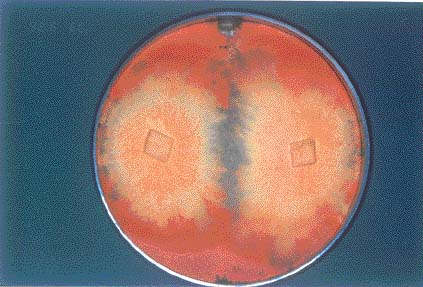
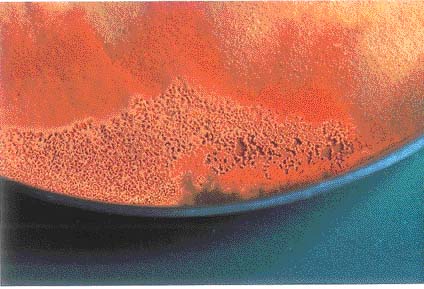
Basidiomatal formation of Antrodia cinnamomea on artificial agar media
Tun-Tschu CHANG* and Wu-Rong WANG
Division on Forest Protection, Taiwan Forestry Research Institute, 53 Nan-Hai Road, Taipei 100, Taiwan
(Received July 19, 2004; Accepted November 2, 2004)
Abstract. Basidiomes of Antrodia cinnamomea are used as a traditional medicine in Taiwan. Because fungal basidiomes are so rare and attempts to cultivate them have failed, the price they command in the marketplace is exorbitant. Here, one dikaryotic isolate from a pairing between two compatible monokaryons obtained from two different basidiomes was able to produce basidiomes on PDA and MEA media at temperatures of 20-28°C within 45 days of incubation. The basidia and basidiospores produced on the hymenial layer were microscopically observed.
Keywords: Antrodia cinnamomea; Basidiomatal formation; Nutrient agar media.
Introduction
Antrodia cinnamomea T.T. Chang & W.N. Chou is a resupinate to effused-reflexed basidiomycete with porous hymenium. It causes wood brown rot of Cinnamomum kanehirai Hayata (Chang and Chou, 1995). Antrodia camphorata S.H. Wu, Ryvarden & T.T Chang has been used for this fungus in the literature but should be avoided because its type material is composed of two different fungal components. Antrodia camphorata is thus considered a nomen confusum with an uncertain nomenclatural status (Chang and Chou, 2004). The fungus is known only from Taiwan and is restricted to C. kanehirai. The basidiomes produced on the infested wood have long been used as a herbal medicine in Taiwan. Owing to its host specificity and rarity in nature as well as effectiveness in curing certain illnesses (Shen et al., 2004), the basidiomes of the fungus are priced high. In the field, the basidiomes grow in the empty rotten trunks of living C. kanehirai and are hardly noticeable until the trees fall down. In order to harvest the basidiomes of A. cinnamomea more easily, some people illegally fell trees of C. kanehirai, an endemic and endangered species to Taiwan, and thereafter, periodically revisit the logging sites to collect the basidiomes. The illegal felling of C. kanehirai in natural forests has severely threatened C. kanehirai. The artificial cultivation of A. cinnamomea basidiomes to satisfy market demand is considered the most effective solution. Although mycelia of A. cinnamomea are easily cultured in/on artificial media, there are no convincing reports on basidiomatal production in pure cultures. Also, the cultured mycelia did not produce certain specific triterpenoids such as zhankuic acids A, B, C, or antcin K, which were only isolated from
basidiomes of A. cinnamomea and are considered to be effective compounds for curing certain illnesses (Shen et al., 2004). In this paper, the basidiomatal formation of A. cinnamomea on artificial agar media is reported, and its teleomorphic structures were also observed to indicate the completion of the life cycle.
Materials and Methods
Fungal Isolates
Basidiomes and basidiospores of A. cinnamomea from two collections (TFRI B496 from Tona, Kaohsiung and TFRI B502 from Alishan, Chiai) were used for isolating dikaryons and monokaryons, respectively. The cultures obtained from the basidiomes of the two collections were dikaryons. The methodology of Chang and Chou (2004) was followed for the isolation of monokaryons. Pieces of pore surface cut from fresh basidiomes were placed into a test tube containing sterile distilled water, and mechanically stirred for 5 min to obtain a basidiospore suspension. The basidiospore suspension was spread onto modified MEA (0.1% malt extract, 2% glucose, and 2% Bacto agar) plates, and colonies resulting from single basidiospores were transferred to MEA (2% malt extract, 2% glucose, and 2% Bacto agar) as monokaryons after incubation at 24°C for 7-10 days. The mated dikaryons obtained from pairings were made by placing mycelial blocks (2 mm in diam.) of two compatible monokaryons at a distance of 1 cm on MEA plates (Chang and Chou, 2004).
Basidiomatal Formation on Artificial Media
Two natural dikaryotic isolates (TFRI B496 and TFRI B502) and 128 mated dikaryotic isolates, obtained from pairings among monokaryons isolated from basidiomes of TFRI B496 and TFRI B502 collections, were used for test
*Corresponding author. E-mail: ttchang@serv.tfri.gov.tw