Bot. Bull. Acad. Sin. (2005) 46: 155-162
LI and XIA Genetic variation of Glyptostrobus pensilis
Population structure and genetic diversity of an endangered species, Glyptostrobus pensilis (Cupressaceae)
Fagen LI and Nianhe XIA*
South China Botanical Garden, The Chinese Academy of Sciences, Guangzhou 510650, P.R. China
(Received January 2, 2004; Accepted December 14, 2004)
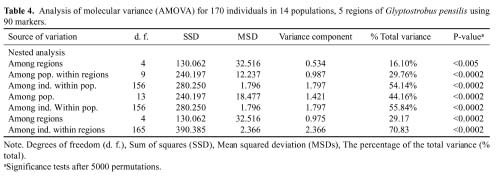
Abstract. Glyptostrobus pensilis (Staunton ex D. Don) K. Koch is a critically endangered species, only distributed in South China and Vietnam. Genetic variation within and between populations was investigated using inter-simple sequence repeat (ISSR). One hundred and seventy individuals from fourteen populations representing five regions in southern and southeastern China were sampled. The results show that genetic diversity of G. pensilis is rather low: P = 10.2%, Ae = 1.070 and Hpop = 0.058 at population level; P = 24.7%, Ae = 1.140 and Hsp = 0.122 at species level. The results further indicate significant genetic differentiation between regions (16.10%, analysis of molecular variance [AMOVA], p<0.005), between populations within particular regions (29.76%, P<0.0002), and between individuals within populations (54.14%, P<0.0002). High levels of genetic differentiation between populations were also detected based on Nei's genetic diversity analysis (51.36%), and Shannon's diversity index (52.54%). Possible reasons for the high genetic differentiation and its low level of diversity within populations are discussed. Possible in situ and ex situ conservation strategies for G. pensilis are suggested.
Keywords: Conservation strategies; Endangered species; Genetic diversity; Glyptostrobus pensilis; ISSR (inter-simple sequence repeat).
Introduction
Glyptostrobus pensilis (Staunton ex D. Don) K. Koch has been recognized as a unique monotypic genus in the family Cupressaceae on the basis of its distinct morphology (Zheng and Fu, 1978; Farjon and Page, 1999). Fossil evidence indicates that G. pensilis was widely distributed in the Far East, Siberia, Canada, and the United States in the Late Cretaceous, and in Siberia and Thailand in the Pliocene (Florin, 1963; Yu, 1995). Due to the early Pleistocene glaciations and subsequent desertification during the Quaternary, this species occurs now only in South China and Vietnam (Farjon and Page, 1999; Li and Xia, 2004). Glyptostrobus pensilis is a monoecious, wind-pollinated, heliophilous and sub-evergreen tree, which is tolerant of waterlogging and can grow in most soils that are not saline or alkaline. The species is adapted to warm and humid habitats. Seedlings or saplings demand sufficient sunlight in addition to fertile and moist soil (Fu and Jin, 1992; Yu, 1995). Glyptostrobus pensilis has been classified as endangered by the IUCN categories and listed in the China Plant Red Data Book: Rare and Endangered Plants (Fu and Jin, 1992; Farjon and Page, 1999). Previous studies of G. pensilis focused mostly on chemical composition (Lan et al., 1992), cytology (Xiao and Dong, 1983; Li, 1987), pollen morphology (Xi, 1986), ecology (Yu, 1995),
and geography (Xu and Li, 1959). Research on the genetic variation within and among populations of G. pensilis has not been reported to date. Genetic variation within and among populations of rare and endangered species plays a significant role in the formulation of appropriate management strategies directed towards their conservation (Milligan et al., 1994).
In recent years, a relatively novel molecular technique that permits the detection of polymorphisms in micro-satellite and inter-micro-satellite loci without previous knowledge of the DNA sequences has been described: "inter-simple sequence repeat PCR" (ISSR-PCR) (Zietkiewicz et al., 1994). This technique has been widely used to investigate genetic diversity and population genetic structure because it overcomes some limitations of allozymes and requires a small amount of DNA (Wolfe et al., 1998; Ratnaparkhe et al., 1998; Esselman et al., 1999). Each primer is composed of di- or trinucleotide repeats with or without one to three additional nucleotides at the 5´- or 3´- end to serve as anchors against reverting strand slippage during amplification (Gupta et al., 1994). The sequences of repeats and anchor nucleotides are arbitrarily selected, and identical repeat sequences with different anchoring sequences can yield different banding patterns (Wolfe et al., 1998). In contrast with other molecular techniques, the sequences that ISSR target are abundant throughout the eukaryotic genome and evolve rapidly. Consequently ISSR may reveal a much higher number of polymorphic fragments per primer than random amplified
*Corresponding author. Tel: + 86-20-37252565; Fax: + 86-20-37252565; E-mail: nhxia@scbg.ac.cn