Bot. Bull. Acad. Sin. (2005) 46: 175-182
CHEN et al. RNAi expression for chloroplastic SOD gene
Antisense and RNAi expression for a chloroplastic superoxide dismutase gene in transgenic plants
Shu-Yan CHEN, Sa XIAO, Man-Xiao ZHANG, Tuo CHEN, Hui-Chun WANG, and Li-Zhe AN*
School of Life Sciences, Lanzhou University, Lanzhou 730000, Gansu, People's Republic of China
(Received June 16, 2004; Accepted Feburary 22, 2005)
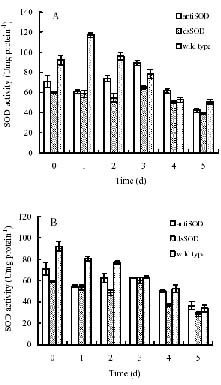
Abstract. The cDNA of tomato chloroplastic Cu/Zn-superoxide dismutase was used to construct transgenic tobacco (Nicotiana plumbaginifolia). It was found that the gene expression of Cu/Zn-superoxide dismutase can constitutively be reduced in the transgenic tobacco plants because of double-stranded (dsRNA) expressed in the form of intron-spliced hairpin structures and antisense suppression. Furthermore, the endogenous chloroplastic Cu/Zn-superoxide dismutase gene in tobacco was established as a target for silencing due to the operation of hpRNA and antisense RNA constructs. However, under salt and PEG-induced stress, the cytosolic Cu/Zn-superoxide dismutase activity in transformed plants obviously increased. At the same time, the PEG pretreatment was able to promote tolerance of the transgenic plants to the salt stress. These results indicated that the use of hpRNA and anti-sense was able to successfully knockout the transcript encoding a chloroplast superoxidase dismutase. The null transformed plants grown under stress produced or retained other superoxide dismutase to compensate for the loss of the chloroplast. Consequently, hpRNA constructs would be helpful in discovering and validating the endogenous chloroplastic Cu/Zn-superoxide dismutase gene, and to prove the cytosolic alternative pathway of plant antioxidation associated with the function of Cu/Zn-superoxide dismutase gene under an unfavorable environment. Meanwhile, the experiment will provide an important technique for the antisensing strategies operating in tobacco.
Keywords: Double-stranded RNA; Salt stress; Superoxide dismutases; Transgenic tobacco.
Introduction
Double-stranded RNA (dsRNA)-mediated interference with expression of specific genes has been observed in a number of organisms including nematodes (Fire et al., 1998), Drosophila (Kennerdel and Carthew, 1998) and plants (Chuang and Meyerowitz, 2000). Such self-complementary hairpin RNA (hpRNA, the expression form of dsRNA in plants) can elicit a high degree and frequency of posttranscriptional gene silencing (PTGS) of endogenous genes (Chuang and Meyerowitz, 2000; Wesley et al., 2001) and has great potential as a tool for gene discovery and validation (Wesley et al., 2001).
In natural conditions plants are often exposed to various environmental stresses that can decrease production. Like other biotic, abiotic, and xenobiotic stresses, salt and drought stresses produce many degenerative reactions mediated by toxic reactive oxygen species (ROS) such as superoxide (O2-), hydrogen peroxide (H2O2), singlet oxygen, and hydroxyl radical, produced when the H2O2 reacts with reduced metal irons such as Fe. These ROS are cytotoxic and can seriously disrupt normal metabolism by way of oxidative damage to lipids (Wise and Naylor, 1987), nucleic acids (Imlay and Linn, 1988), and proteins (Davis, 1987), resulting in mutation, protein destruction, and peroxidation of membrane lipids respectively, which in turn may lead to
diseases and degenerative processes (carcinogenesis and immunodeficiency, for example). There is circumstantial evidence that high levels of SOD activity in plants result in tolerance of a variety of environmental and chemical challenges and that overexpression of SOD acts as a safeguard against both drought (Mckersie et al., 1996; Bowler et al., 1992) and salinity (Zhu, 2002; Hasegawa et al., 2000). On the other hand, although transgenic tobacco plants could express high levels of petunia chloroplastic Cu/Zn-SOD (Tepperman and Dunsmuir, 1990), no detectable increase in the protection of photosynthetic activity was detected in these plants after their exposure to MV, nor was there any reduction in symptom development after ozone fumigation (Pitcher et al., 1991).
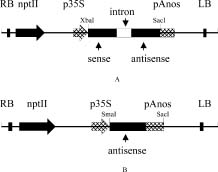
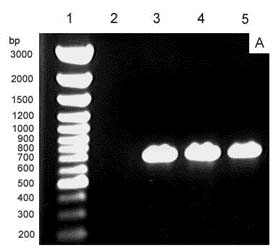
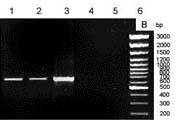
Although many previous reports have affirmed the protective capabilities of SOD as an antioxidant enzyme and its role in cross tolerance for many stresses, much remains to be clarified, especially the role of SOD isozymes in different cell compartments and in the cross-tolerance mechanism. In this study, we made use of the cDNA of tomato (Lycopersicon esculentum cv E.C.) chloroplastic Cu/Zn-SOD to construct transgenic tobacco (Nicotiana plumbaginifolia). The result showed that this can constitutively reduce chloroplastic Cu/Zn-SOD gene expression in tobacco through hpRNA and antisense suppression. To the best of our knowledge, this is first report that demonstrates the expression of the Cu/Zn-SOD gene in the chloroplast of transgenic tobacco to be repressed, while it is increased in the cytosol for protection against salinity (NaCl) and polyethylene glycol-induced (PEG) stress. Can
*Corresponding author. Tel: +86-931-4967181; Fax: +86-931-8912561; E-mail: csy522@sina.com.cn; lizhean@lzu.edu.cn