Bot. Bull. Acad. Sin. (2005) 46: 189-196
KOKILADEVI et al. Characterization of a-amylase inhibitor in Vigna sublobata
Characterization of Alpha-amylase inhibitor in Vigna sublobata
E. KOKILADEVI*, A. MANICKAM, and B. THAYUMANAVAN
Center for Plant Molecular Biology, Tamil Nadu Agricultural University, Coimbatore 641 003, Tamil Nadu, India
(Received August 30, 2004; Accepted January 14, 2005)
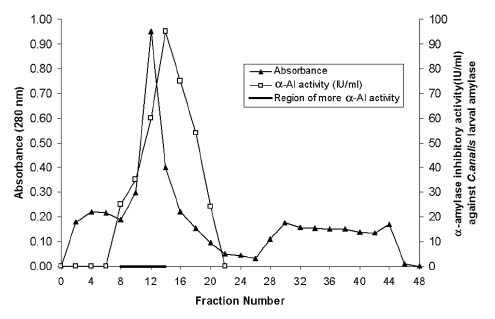
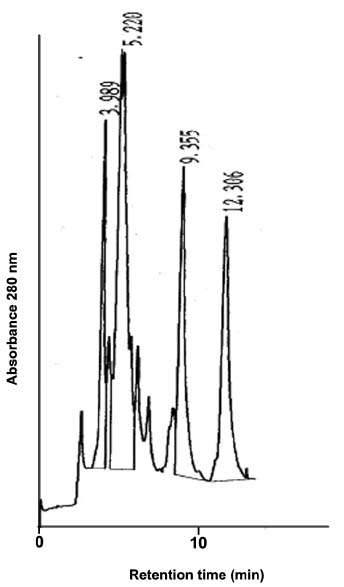
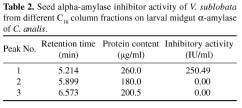
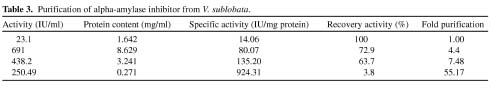
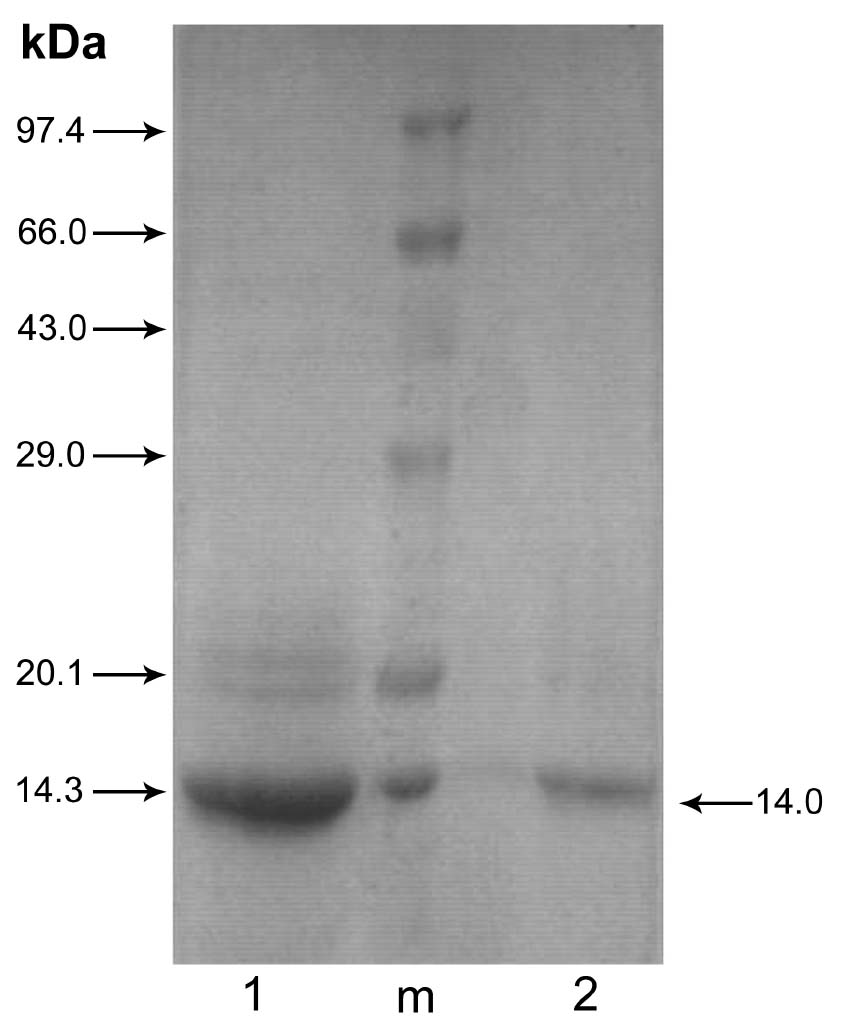
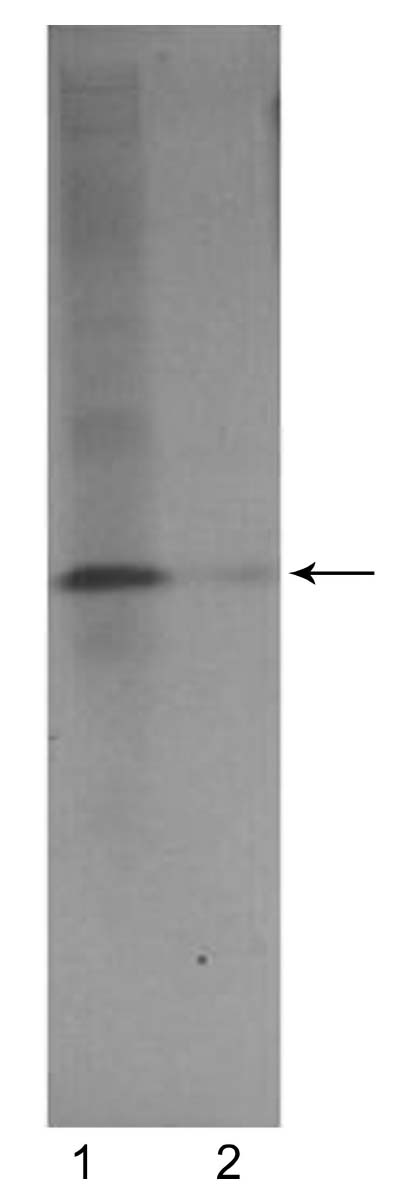
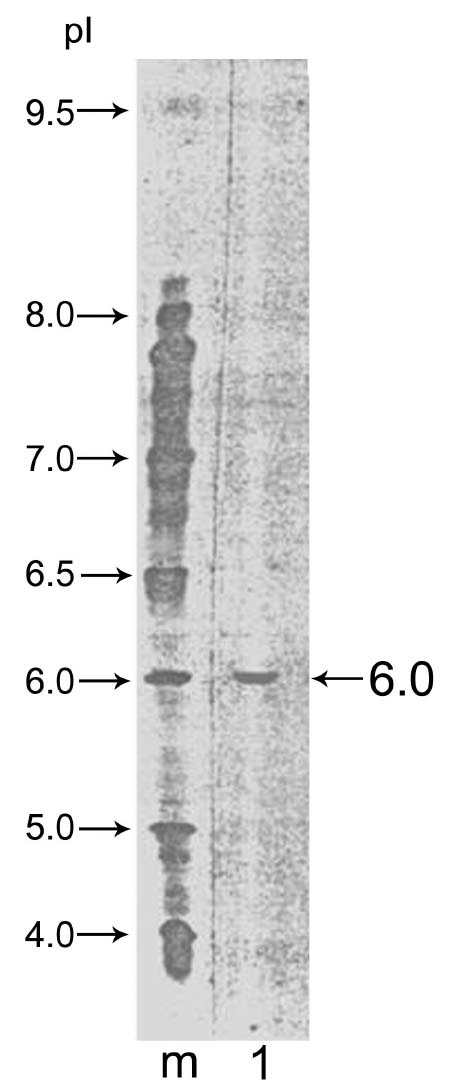
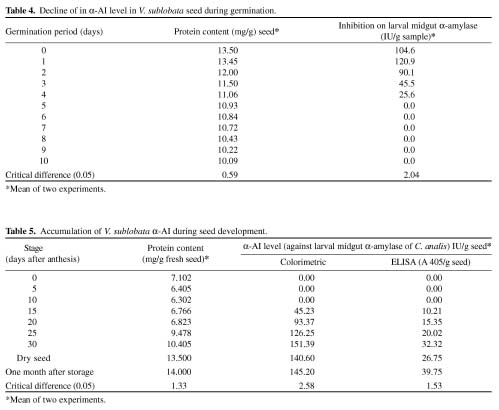
Abstract. Alpha-amylase inhibitor protein, which inhibits the activity of insect (Callosobruchus analis) a-amylase, was characterized from V. sublobata. The molecular weight of purified inhibitor protein was 14 kDa by SDS-PAGE. The inhibitor is non-glycosylated protein and its N-terminal sequence is similar (A P S P V ) to Phaseolus vulgaris a-AI-1. Its pI value is 6.0 and largely localised in cotyledons. The inhibitory activity decreased during germination from days one to five. In the developmental stages of seed formation from anthesis to 30 days the inhibitor content increased.
Keywords: a-Amylase; Alpha-amylase inhibitor; Callosobruchus analis; Vigna sublobata.
Introduction
Many edible plant seeds contain substances that inhibit enzymes, especially hydrolases. Most of these compounds are proteins by nature, which specifically inhibit enzymes by forming complexes that block the active site or alter enzyme conformation, ultimately reducing the catalytic function. Substances present in the seeds of P. vulgaris suggested to play a role in insect resistance include heteropolysaccharides, lectins, and protease- and amylase inhibitors (Applebaum et al., 1969). It has also been suggested that the various forms of resistance are mainly due to the presence of genes of the lectin family (Chrispeels and Raikhel, 1991).
Among the storage pests, the genus- Callosobruchus (Coleoptera: Bruchidae) is the most serious pest on mungbean and other Vigna species (Talekar and Lin, 1981). Alpha-amylase inhibitors have been studied in several common beans (Marshall and Lauda, 1975) maize (Blanco-Labra and Iturbe-Chinas, 1981), sorghum (Kutty and Pattabiraman, 1986), wheat (Warchalewski, 1977), and barley (Mundy and Rogers, 1986). Seed alpha-amylase inhibitor (a-AI) in several cultivars of the common bean plays a protective role against bruchid pests (Ishimoto and Kitamura, 1989). The a-AI strongly inhibited the larval midgut a-amylase activities of adzuki bean weevil (C. chinensis L.) and cowpea weevil (C. maculatus), non-pest species of the common bean. The gene encoding an a-AI was isolated from common bean and transferred to pea (Pisum sativum), and transgenic peas were produced that expressed the transgene (Shade et al., 1994). Bean a-AI-1 in transgenic peas provided complete protection from pea weevil (Bruchus pisorum) under field conditions (Morton et al., 2000). Earlier studies from our laboratory also re
ported that the presence of a-AI is one among the possible strong factors for bruchid resistance in the wild relatives of mungbean, V. sublobata (Sahu, 1996). In the present study an a-AI from V. sublobata has been isolated from seed and characterized.
Materials and Methods
Source of Seed Materials
Seeds of Vigna genotypes, including viz., green gram (var. CO 4, CO 5, K-851, P-9078), black gram (var. CO 5), V. glabracens, V. sublobata, V. trilobata and V. umbellata obtained from the Department of Pulses, Center for Plant Breeding and Genetics, Tamil Nadu Agricultural University, Coimbatore were used.
Extraction of Seed Proteins
The extraction of proteins from seed flour was done following the method of Mirkov et al. (1994) for electrophoresis (SDS-PAGE) and immunological analysis. One hundred mg of finely ground seed flour was extracted in the extraction buffer (10 mM Tris-HCl [pH 7.5], 500 mM NaCl, 1% 2-mercapto-ethanol, 0.1% triton-X-100, 2 mM phenylmethylsulphonyl fluoride [PMSF] [1 ml/mg sample]) by homogenization followed by incubation at 4°C for 1 h. It was then centrifuged at 15,000 rpm for 15 min at 4°C. The supernatant was collected and stored frozen in aliquots. The protein content in the extract was quantified following Bradford's (1976) method.
a-AI Assay
The a-AI was assayed by quantifying the reducing sugar (maltose equivalent) liberated under the assay conditions. The enzyme inhibitory activity is expressed as the decrease in units of maltose liberated. A modified dinitrosalicylic acid (DNS) method of Bernfeld (1955) was
*Corresponding author. E-mail: cmkokila@yahoo.com