Bot. Bull. Acad. Sin. (2005) 46: 197-203
LIU et al. Effect of ulvoid algae on Thalassia hemprichii
The effect of ulvoid macroalgae on the inorganic carbon utilization by an intertidal seagrass Thalassia hemprichii
Shao-Lun LIU1, Wei-Lung WANG1,*, Danilo T. DY2, and Cheng-Chang FU3
1Department of Biology, National Changhua University of Education, Changhua 500, Taiwan
2Marine Biology Section, Department of Biology, University of San Carlos, Cebu City 6000, Philippines
3Oldinburgh Company, Ltd., Taipei 100, Taiwan
(Received October 12, 2004; Accepted January 27, 2005)
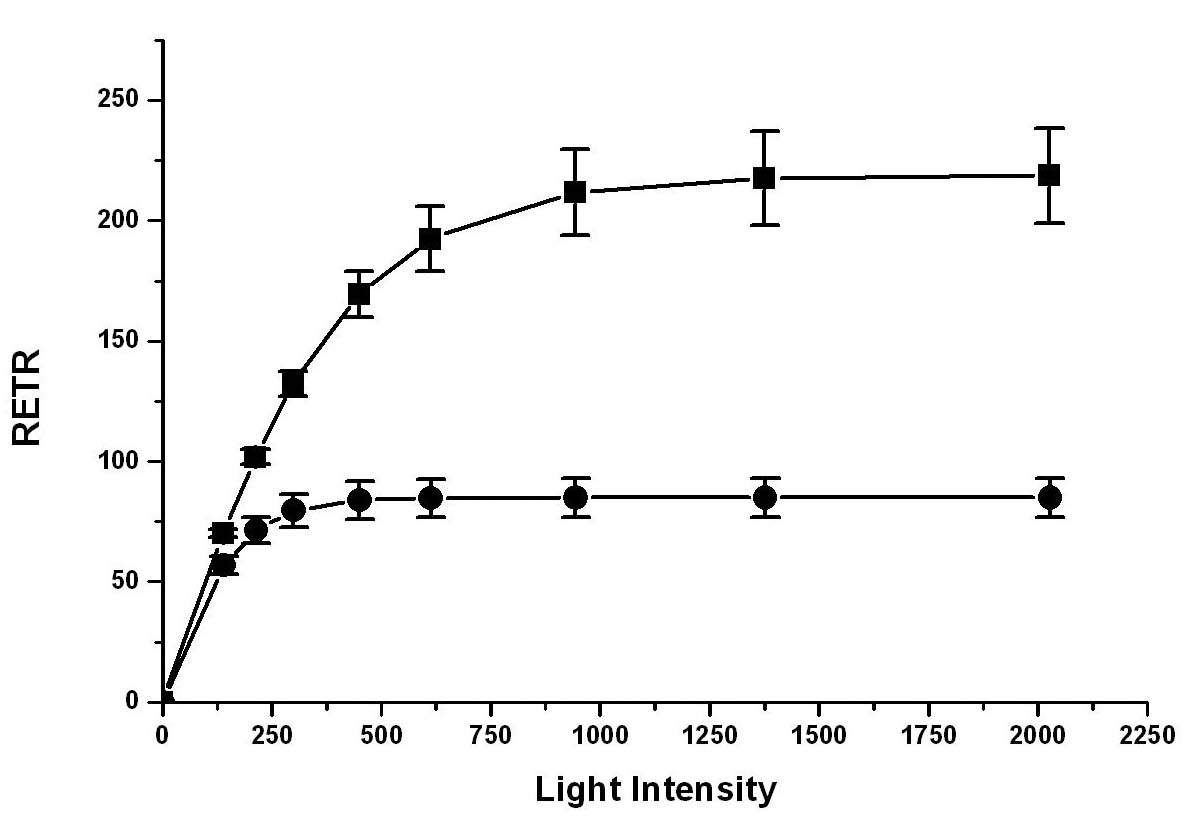
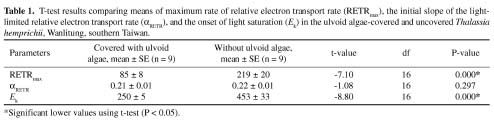
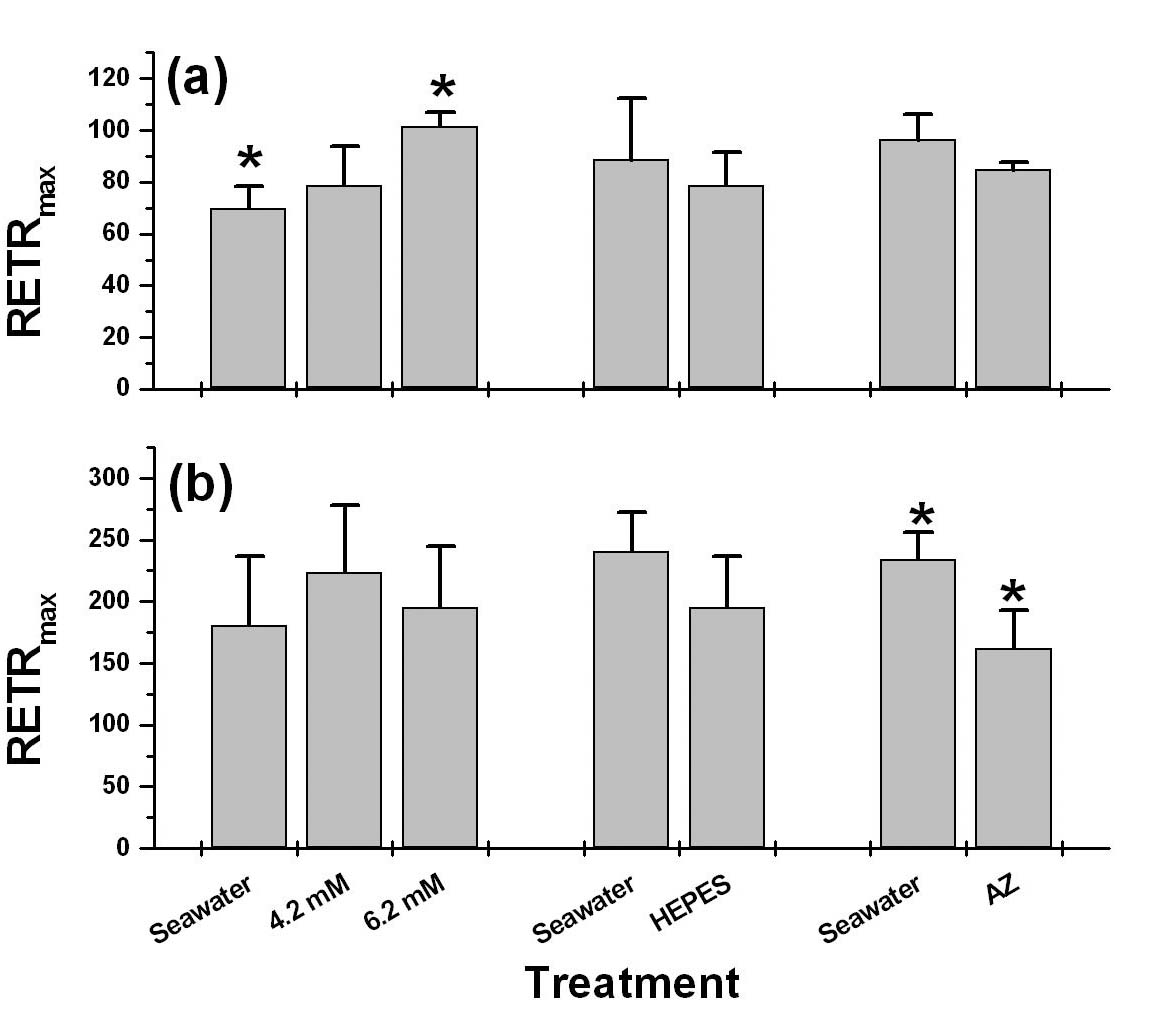
Abstract. Blooms of ulvoid macroalgae (mainly Enteromorpha and Ulva) have covered 80% of the intertidal seagrass bed at Wanlitung, southern Taiwan, effectively shading the seagrasss species Thalassia hemprichii resulting in a decrease in photosynthetic performance and low inorganic carbon (Ci) uptake. We looked for evidence of Ci limitation and investigated the Ci utilization characteristics of ulvoid-free and ulvoid-covered T. hemprichii. The rapid light curve (RLC) function of the Diving-PAM (Diving-PAM, Walz, Germany) was used to measure in situ photosynthetic performance (based on the effective quantum yield of PSII [Y] values) of intact seagrasses that were placed in small incubating chambers. Significantly, a lower RETRmax (maximum relative electron transport rate) and Ek (light intensity at the onset of saturation) were noted in the ulvoid-covered compared to the ulvoid-free T. hemprichii, suggesting that the former has acclimatized to the low light environment becoming a "shade type" plant. The ulvoid-covered T. hemprichii showed some evidence of Ci limitation since a significant increase in RETRmax (up to 46%; P < 0.05) was noted after an increase in the concentration of NaHCO3 from 2.2 (normal seawater) to 6.2 mM. In terms of Ci utilization characteristics, T. hemprichii could directly absorb HCO3- as the major Ci source but partially depended on the extracellular carbonic anhydrase (CA) to convert HCO3- to CO2 prior to uptake in the ulvoid-free, high light-adapted populations. A wastewater stream with a high nutrient load coming from the urbanized area may have caused the frequent blooms of ulvoid macroalgae.
Keywords: Bicarbonate; Photosynthesis; Taiwan; Ulvoid bloom.
Abbreviations: AF, absorption factor; AZ, acetazolamide; CA, carbonic anhydrase; Ci, inorganic carbon; DIN, dissolved inorganic nitrogen; DIP, dissolved inorganic phosphate; Ek, the light intensity at the onset of photosynthetic saturation; RETR, relative electron transport rate; HEPES, N-[2-Hydroxyethyl]piperazine-N'-[2-ethanesulfonic acid]; F, minimal chlorophyll-a fluorescence in the light-adapted state; Fm', maximum chlorophyll-a fluorescence in the light-adapted state; PAM, pulse amplitude modulated; PPFD, photosynthetic photon flux density; RETRmax, maximum relative electron transport rate; PSII, photosystem II; Y, the effective quantum yield of PSII; aRETR, the initial slope of the light-limited relative electron transport rate.
Introduction
Seagrasses are submerged marine angiosperms that comprise about 60 species worldwide (Den Hartog, 1970; Green and Short, 2003), seven of which have been recorded from Taiwan (Yang et al., 2002). To date, very few studies on seagrasses in Taiwan have appeared (Mok et al., 1993; Shieh and Yang, 1997; Lin and Shao, 1998; Yang et al., 2002). Lin and Shao (1998) reported that Thalassia hemprichii (Ehrenberg) Ascherson from southern Taiwan was threatened by anthropogenic activities due to excessive nutrient input. The nutrient over-enrichment of coastal waters can lead to the proliferation of bloom-forming "ephemeral" macroalgae which end up shading seagrass populations and eventually displacing seagrass as the dominant benthic autotrophs (McGlathery, 2001).
Manipulative experiments on a naturally occurring bloom of Ulvaria obscura (Kützing) Gayral in Armitage Bay, Blakely Island, Washington State, USA resulted in reduced seagrass shoot density (Nelson and Lee, 2001). Competition with other macrophytes for one or more resources, such as light, O2, or inorganic carbon (Ci) could be behind the seagrass growth reduction (Coffaro and Bocci, 1997) suggesting some form of resource limitation.
At Wanlitung (N 21º99'60", E 120º70'50'') in southern Taiwan frequent blooms of ulvoid macroalgae (mainly Enteromorpha spp. and Ulva spp.) were observed in the intertidal habitat covering some portion of the flourishing seagrass, Thalassia hemprichii. A continuous wastewater stream from the urbanized area was observed to drain into the seagrass habitat. Dissolved inorganic nitrogen (DIN) was present at 0.51 ppm during low tide and 0.49 ppm during high tide. Dissolved inorganic phosphorus (DIP) was 0.24 ppm during low tide and 0.03 ppm during high tide (Liu and Wang, unpublished data). The DIN:DIP
*Corresponding author. E-mail: wlwang@cc.ncue.edu.tw