Bot. Bull. Acad. Sin. (2005) 46: 217-222
WANG et al. Nuclear transplantation in Antrodia cinnamomea
Variation and segregation following nuclear transplantation in Antrodia cinnamomea
Wei-Ming WANG1, Rey-Yuh WU1, and Wen-Hsiung KO2,*
1Development Center for Biotechnology, 101 Lane 169, Kang Ning St., Hsi Chih, Taipei, Taiwan
2Department of Plant and Environmental Protection Sciences, University of Hawaii at Manoa, Hilo, Hawaii 96720, USA
(Received July 29, 2004; Accepted January 27, 2005)
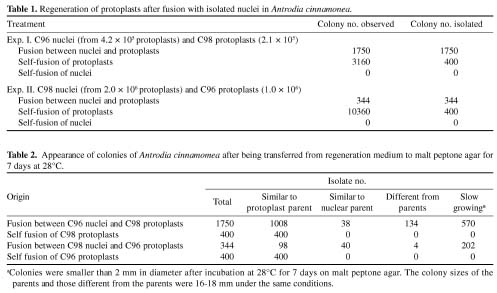
Abstract. Intraspecific fusion of protoplasts with nuclei in Antrodia cinnamomea lengthened the time for development of colonies on regeneration medium and reduced the number of protoplasts capable of regeneration. When 2.1 × 105 C98 protoplasts were fused with C96 nuclei, 1750 colonies appeared on the regeneration medium, and 742 of them were putative nuclear hybrids as their colonies were different from that of the protoplast parent. Without the presence of C96 nuclei, 3160 colonies appeared on the regeneration medium and 400 of them tested were similar to the protoplast parent in colony appearance. No colonies appeared when self-fused nuclei were plated on regeneration medium. The rate of successful transfer was 23%. The success rate was 2.4% for reciprocal transfer. About 77 to 82% of nuclear hybrids were slow in growth. These isolates resumed normal growth upon subculture. All the nuclear hybrids obtained were unstable. Segregation occurred during growth of mycelia, resulting in the formation of colonies different from those of the originals in subcultures. After five consecutive subcultures, 13 stable nuclear hybrids were obtained. These isolates differed greatly from the parental isolates and from each other in pigmentation, growth rate, and colony pattern.
Keywords: Antrodia cinnamomea; Colony morphology; Nuclear hybrid; Nuclear transplantation.
Introduction
Antrodia cinnamomea T. T. Chang & W. N. Chou is a rare and extremely valuable medicinal fungus in Taiwan (Chang and Chou, 1995, 2004). The basidiomes of the fungus are well known locally for their effectiveness in treating various human illnesses, including liver cancer, kidney problems, diabetes, and food poisoning (Chen et al., 2001b). New compounds with different medicinal properties have been isolated from these basidiomes in recent years (Chen and Yang, 1995; Cherng and Chiang, 1995; Cherng et al., 1995; Chiang et al., 1995). Antrodia cinnamomea is indigenous to Taiwan, and the protected precious endemic tree species Cinnamomum kanehirai Hayata is its only known host (Chang and Chou, 1995). Since it is still impossible to produce fruiting bodies of A. cinnamomea in cultures, the basidiomes of the fungus collected from natural forests by the natives are in great demand and have been sold for about U.S. $15,000 per kg (Wu et al., 1997). Some medicinal effects have been detected in liquid as well as solid cultures of A. cinnamomea (Chen et al., 2001a; 2001b). Isolates of this fungus with the ability to produce large quantities of medicinally ac
tive compounds are, therefore, urgently needed. Unfortunately, the number of isolates available for selection is very limited because of the plant's rarity in nature.
Recently, hybrids of Phytophthora resulting from nuclear transplantation have been shown to display increased variation in both physiological and morphological characteristics (Gu and Ko, 1998; 2000; 2001). A project was, therefore, initiated to develop a protocol for the nuclear transplantation of A. cinnamomea to generate variation for selection of strains capable of producing large quantities of medicinal compounds. Reports on nuclear transplantation in fungi are few. Successful application of nuclear transplantation requires a selective method for isolation of nuclear hybrids. Initially, complementary auxotrophic mutants were used, and prototrophic strains capable of growing on minimal medium subsequent to nuclear transplantation were selected as nuclear hybrids (Ferency and Pesti, 1982: Sivan et al., 1990; Vagvolgyi and Ferenczy, 1992). Recently, two mutants, each resistant to a different fungicide, were used, and strains capable of growing on medium containing both fungicides were selected as nuclear hybrids (Gu and Ko, 1998; 2000; 2001). The generation of auxotrophic mutants and induction of fungicide resistant mutants are tedious and time consuming tasks. In this study we developed a method for detection of nuclear hybrids based on a comparison of colony morphology with parental types.
*Corresponding author. Tel: +1-808-974-4105; Fax: +1-808-974-4110; E-mail: kowh@hawaii.edu