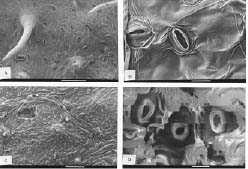
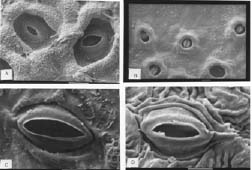
Bot. Bull. Acad. Sin. (2005) 46: 231-238
SONIBARE et al. Scanning electron microscopy studies of Ficus
A survey of epidermal morphology in Ficus Linn. (Moraceae) of Nigeria
Mubo A. SONIBARE1,*, Adeniyi A. JAYEOLA2, Adeyemi EGUNYOMI2, and Jin MURATA1
1Koishikawa Botanical Gardens, Graduate School of Science, Faculty of Science, University of Tokyo, Japan
2Department of Botany and Microbiology, University of Ibadan, Nigeria
(Received July 14, 2004; Accepted January 14, 2005)
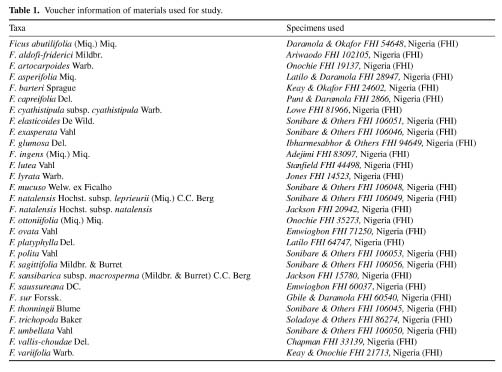
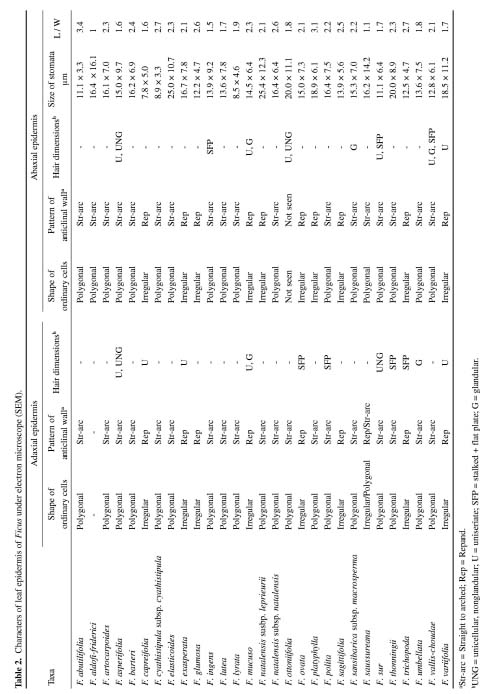
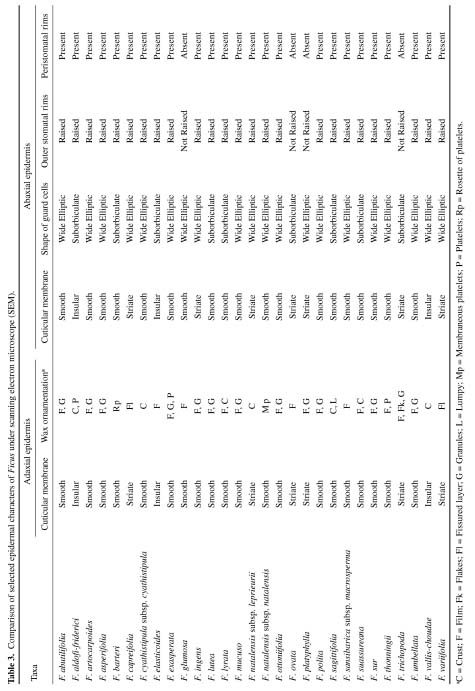
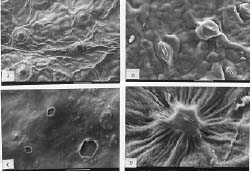
Abstract. A detailed morphological study of the leaf epidermis of the tropical genus Ficus Linn. in Nigeria is presented. The study revealed several interesting epidermal features that have not previously been reported in the genus. Leaf epidermal characters such as pattern of epidermal cells, type of stomata, shape of guard cell pairs, and cuticular ornamentation are constant in some species and variable in others and thus of great significance in understanding the relationships between and within species. Leaves are hypostomatic in all species except in F. vallis-choudae Del., which has amphistomatic leaves. Poral epidermal walls of the outer stomatal ledge are distinctly higher than the peristomatal rims in F. cyathistipula subsp. cyathistipula Warb., F. lyrata Warb., and F. sagittifolia Mildbr. & Burret.
Keywords: Epidermal characters; Ficus; Morphological survey; Scanning electron microscopy (SEM); Taxonomy.
Introduction
Literature on the epidermal morphology of Ficus is relatively rare though the taxonomic value of epidermal morphology is well documented in botanical literature for several other groups of angiosperms (Stace, 1965). As an example, epidermal characters of Euphorbiaceae have been studied by Amelunxen et al. (1967), Kakkar and Paliwal (1974), and Raju and Rao (1977). Several reports were published on the epidermal structures and stomatal ontogeny by Karatela and Gill (1984a, b; 1985) who stressed their usefulness in plant taxonomy.
In his taxonomic study of Jatropha L. (Euphorbiaceae), Dehgan (1980) reported that all species, with the exception of J. fremontioides Standley, have paracytic stomata that are with few exceptions restricted to the abaxial surface. Wax morphology has been used extensively in taxonomic work. The classic work of Hallam and Chambers (1970) on SEM studies of the leaf of Eucalyptus L'Heritier has demonstrated the potential of epicuticular wax studies to aid the classification of a complex genus. Although morphological studies of the epidermis may not themselves be sufficient as taxonomic evidence, they could in conjunction with anatomical features serve as good taxonomic tools for delimiting taxa.
Stomata and associated epidermal cells are providing an increasingly important source of taxonomic characters. The taxonomic significance of stomata distribution and mor
phology in the Epacridaceae was surveyed by Watson (1962), who found that members of the tribe Styphelieae typically possess only adaxial stomata (on the sepal) while in the Epacrideae they are usually abaxial. Trichomes and hydathodes are other characters found on the epidermis and other organs of plants that can serve as good taxonomic tools. They have been employed both for classification and identification purposes by many systematists (Theobald et al., 1979; Rollins, 1993; Potgieter and van Wyk, 1999; Dickison, 2000; Batterman and Lammers, 2004). De Bary (1884) and Renner (1907) have also discussed hydathodes in Ficus.
Preliminary field observation of Nigerian Ficus populations revealed conspicuous differences in vegetative and floral features among the taxa. Previous classifications of the genus have been largely based on the systematics of pollinating wasps (Corner, 1958, 1965; Ramirez, 1977). It is imperative therefore to attempt a search for epidermal characters that may be of taxonomic utility. The main objective of this work was to survey the leaf epidermis of Ficus species in Nigeria in search of taxonomic characters that might assist in identification and understanding of the variations among the species.
Materials and Methods
Plant samples were obtained from herbarium specimens or personal collections. Small pieces (c. 7 mm2) of leaf from herbarium materials were dehydrated in an absolute ethanol:acetone series (90% ethanol, 30 min; absolute ethanol 30 min; 50:50 absolute ethanol:acetone, 10 min; and finally two steps of acetone, 10 min each), according to Luckow and Grimmes (1997). Dehydrated material was then critical point dried using an Oriental 100V drier and sputter coated with
*Corresponding author. Present address: Department of Biological Sciences, Olabisi Onabanjo University, P.M.B. 2002, Ago-Iwoye, Nigeria. Tel: 234-803-365-9517; Fax: 234-2-8103043; E-mail: sonibaredeola@yahoo.com