Bot. Bull. Acad. Sin. (2005) 46: 293-299
YANG et al. Seed germination eco-physiology of Mikania micrantha
Seed germination eco-physiology of Mikania micrantha H.B.K
Qi-He YANG1, 2, Wan-Hui YE1,*, Xiong DENG1, Hong-Ling CAO1, Yun ZHANG1, and Kai-Yang XU1
1South China Botanical Garden, the Chinese Academy of Sciences, Guangzhou 510650, The People's Republic of China
2Jiaying College, Meizhou 514015, The People's Republic of China
(Received May 31, 2004; Accepted February 24, 2005)
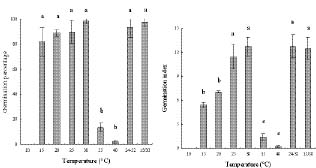
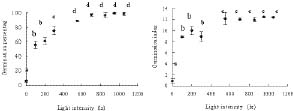
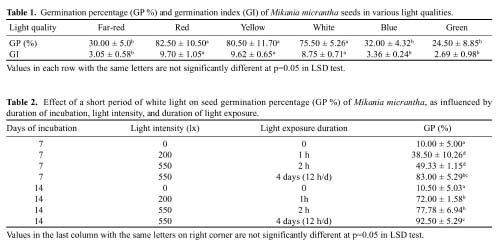
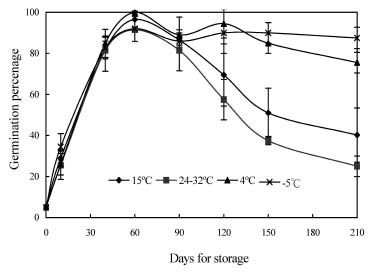
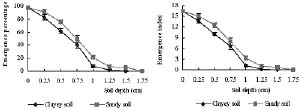
Abstract. Mikania micrantha H.B.K is an invasive alien weed in South China. To help understand the mechanisms of its invasiveness, we studied its seed germination ecophysiology. We found that the optimal temperatures for seed germination were 25, 30, 15/30°C (night/day), and an ambient temperature (24-32°C). Germination percentage (GP) in dark was less than 10%, and it increased with increasing light intensity up to 700 lx. However, GP and germination index (GI) did not change significantly when light was more than 700 and 550 lx, respectively. Germination was enhanced by white, yellow and red light more than far-red, blue, and green light. Light sensitivity and germination of seeds increased with increasing dark incubation time. Newly ripened seeds exhibited innate dormancy that was released by a two-month after-ripening period in dry storage. Except at a storage temperature of -5°C, seed GP decreased when storage time exceeded the after-ripening period, and this decrease was more pronounced from a storage temperature of 4°C to 15°C to ambient 24-32°C. Percentage seedling emergence was lower and slower when seeds were covered deeper in soil, and no emergence occurred when seeds were buried at 1.5 and 1.75 cm in clayey and sandy soil, respectively. With increased soil moisture content (MC) from 8% to about 20%, seedling emergence became increased and quickened. However, an MC greater than 23% reduced emergence. These responses of seed germination to various treatments indicate that this weed is well adapted to environments in South China.
Keywords: Germination; Light; Mikania micrantha, Seed; Soil depth; Soil moisture; Soil type; Temperature.
Introduction
Mikania micrantha (hereafter Mikania), a member of the Asteraceae, originates in Central and South America. In its native land, it is a component of aquatic ecosystems such as marshes and riverbanks and rarely grows in other habitats (Ye and Zhou, 2001). However, it is very variable in form, and in many areas it is extremely invasive. Mikania has been called "Mile-a-minute" and "plant-killer" (Waterhouse, 1994; Ye and Zhou, 2001). The species is widespread throughout tropical Asia, including India, Malaysia, Thailand, and Indonesia, and it has recently been observed in Nepal and Australia. Mikania also occurs in Papua New Guinea, the Solomon Islands, the Philippines, Christmas Island in the Indian Ocean, and on Pacific Ocean islands including Fiji and Western Samoa (Evans et al., 2001). Since the 1980s, it has been found in South China. Its distribution is much wider than previously thought (Sankaran et al., 2001).
Mikania is one of the 100 most serious tropical weeds in the world. It is a fast growing perennial creeping vine that colonizes agricultural land and damages tree crops, agroforestry, and multipurpose trees in moist forest zones of Asia, particularly Southeast Asia (Muniappan and Viraktamath, 1993; Waterhouse, 1994) and South China (Huang et al., 2000). The species causes substantial yield
losses in agroforestry systems, in tea, oil palm, rubber, teak, and sal (Shorea robusta) plantations, as well as in many crops including bamboo, reed, plantains and pineapples. Mikania has also invaded natural evergreen, semi-evergreen, and moist deciduous forests, and it is threatening the biodiversity of these ecosystems. At present, no efficient ways are available to control this aggressive weedy species (Holm et al., 1977; Ye and Zhou, 2001).
Mkania has vigorous vegetative growth and a high sexual reproductive capacity (Swarmy and Ramakrishnan, 1987), but it multiplies mainly by seed (Holm et al., 1977). Optimal conditions for seed germination and emergence are often a reflection of the optimal growth conditions for the entire life cycle of a plant species, and the survival and invasion of plants are associated mainly with the mechanisms of seed germination and emergence (Raejmanek and Richardson, 1996). Seed morphology influences seed dispersal, and environmental factorssuch as temperature, light, soil moisture content, and soil depthaffect seed germination and seedling establishment (Baskin and Baskin, 1989; Benvenuti et al., 2001). Knowledge about these stages of the life cycle of Mikania will help us understand the optimum conditions for its seed germination and seedling establishment, predict its population dynamics, explain how this species spreads to new areas, and enable us to develop methods to control its invasion. However, we do not have such basic information for the species. Thus, we observed the morphological characteristics of Mikania seeds, studied its seed germination in different storage
*Corresponding author. E-mail: why@scib.ac.cn; yangqihe@scib.ac.cn; Tel: (+86) 13710546160.