Bot. Bull. Acad. Sin. (2005) 46: 301-305
AGAR and ALPSOY Antagonistic effect of selenium
Antagonistic effect of selenium against aflatoxin G1 toxicity induced chromosomal aberrations and metabolic activities of two crop plants
Güleray AGAR* and Lokman ALPSOY
Department of Biology, Faculty of Science and Arts, Atatürk University, 25240/Erzurum, TURKEY
(Received November 15, 2004; Accepted April 15, 2005)
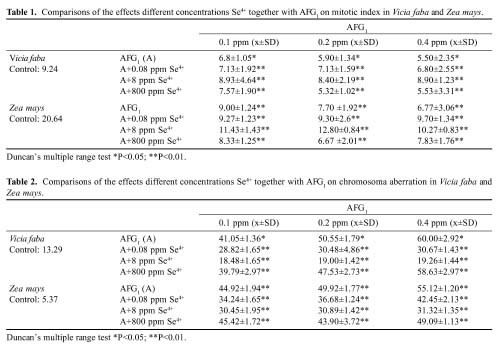
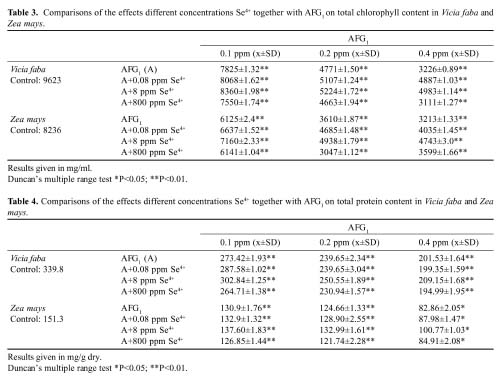
Abstract. The antagonistic effect of selenium (Se4+) against aflatoxin G1 (AFG1) toxicity induced chromosomal aberrations and metabolic activities in plants was studied. The results showed that 0.1, 0.2, 0.4 ppm concentrations of aflatoxin G1 increased chromosomal aberration and mitotic index on total protein, and chlorophyll content decreased. When 0.08, 8, 800 ppm concentrations of Se4+ were added to AFG1, the frequencies of chromosomal aberrations decreased and mitotic index, total protein and chloropyll content increased. Also, results suggested that Se4+ has an antagonistic effect against AFG1, and that the degree of antagonistism of Se4+ against AFG1 is probably related to its concentration ratio.
Keyword: Aflatoxin G1; Genotoxicity; Selenium; Vicia faba; Zea mays.
Introduction
Aflatoxins, the most toxic of all the mycotoxins, have been described as natural contaminants in many consumables. These have been reported to affect the physiological processes of some crops. Most studies done have focused on the effect of aflatoxin B1 (AFB1). Other aflatoxins have received less attention, and very few studies have been carried out with AFG1.
Previous papers on the effect of AFG1 in higher plants indicate that it has an effect on total lipid, protein, and carbohydrate content, chlorophyll synthesis, inhibition of seed germination and growth, and chromosomal aberration (Asahi et al., 1969; Crisian, 1973; Abdou et al., 1984; Abdou et al., 1989; El-Zawahri et al., 1990; El-Naghy et al., 1999).
Recently, several studies have demonstrated that certain dietary substances (such as selenium, b carotene and vitamins) provide a protective effect against AFB1 cytotoxicity in several animal species. The anticarcinogenic and antimutagenic effects of Se4+ against AFB1 have been found in several animal studies (Chen, 1982; Chen et al., 1982; Bhattacharya et al.,1987; Gregory et al., 1984; Bronzetti et al., 2001). However, so far no report has shown a protective effect of Se4+ against AFG1 toxicity. In the present study, we describe the effect of AFG1 on total protein, chlorophyll content, and chromosomal aberration and discuss whether Se4+ has protective effects on these parameters.
Materials and Methods
AFG1 and sodium selenite were obtained from Sigma Chemical Company, USA. Vicia faba seeds were obtained from the Department of Field Crops, Faculty of Agriculture, Atatürk University (Turkey). Zea mays seeds were obtained from the Department of Field Crops, Faculty of Agriculture, Karadeniz Technical University (Turkey). Vicia faba and Zea mays seeds of equal size were chosen and surface sterilized with 2.5 %/w/v of NaOCl for 3 min. After this treatment the seeds were rinsed with four changes of top water, and dried using sterile filter paper. The seeds were soaked in sterile distilled water for 1 h, and then fifteen seeds were germinated in 15 cm diameter petri dishes on four layers of sterile Whatman number 1 filter paper. Solution containing zero (control), 0.1, 0.2, 0.4 ppm of AFG1 and the joint concentration of AFG1 together with 0.8, 8, 800 ppm of Se4+ were added to each plate as seen Tables 1-4. The dishes were allowed to germinate in the dark at 25°C. When the roots reached 1.5-3.0 cm in length, they were collected and fixed in acetic acid: alcohol (1:3).
Cytological preparations were carried out using Feulgen's squash technique. The percentage of mitotic activity (mitotic index-MI.) was calculated by counting the total number of dividing cells to the total number of cells examined. Chromosomal aberrations (CA) were scored at anaphase and telophase, and the mutagenic effect was estimated as the percentage of cells with bridges and fragments. In addition to bridges and fragments, other forms of anomalies were identified in all mitotic stages.
For physiological studies, after 7 days, the total chlorophyll content of 20 g fresh tissue from shoot systems
*Corresponding author. E-mail: gagar@atauni.edu.tr; agar252001@yahoo.com; Tel: +90-442-2314341; Fax: +90-442-2360948.