Bot. Bull. Acad. Sin. (2005) 46: 307-313
ZHANG et al. Photosynthetic performances of transplanted Cypripedium flavum
Photosynthetic performances of transplanted Cypripedium flavum plants
Shi-Bao Zhang1,2, Hong Hu1,*, Zhe-Kun Zhou1, Kun Xu1, Ning Yan1, and Shu-Yun Li1
1Kunming Institute of Botany, The Chinese Academy of Sciences, Kunming 650204, P.R. China
2Graduate School of the Chinese Academy of Sciences, Beijing 100039, P.R. China
(Received September 23, 2004; Accepted May 5, 2005)
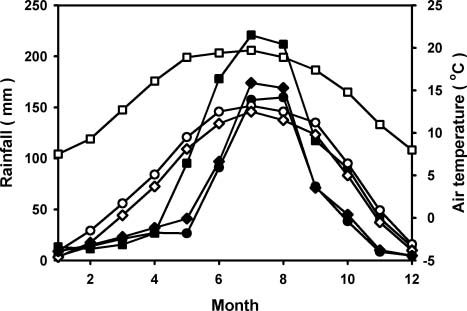
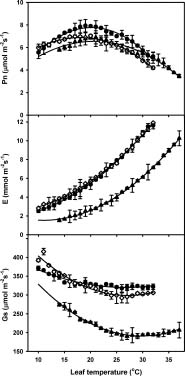
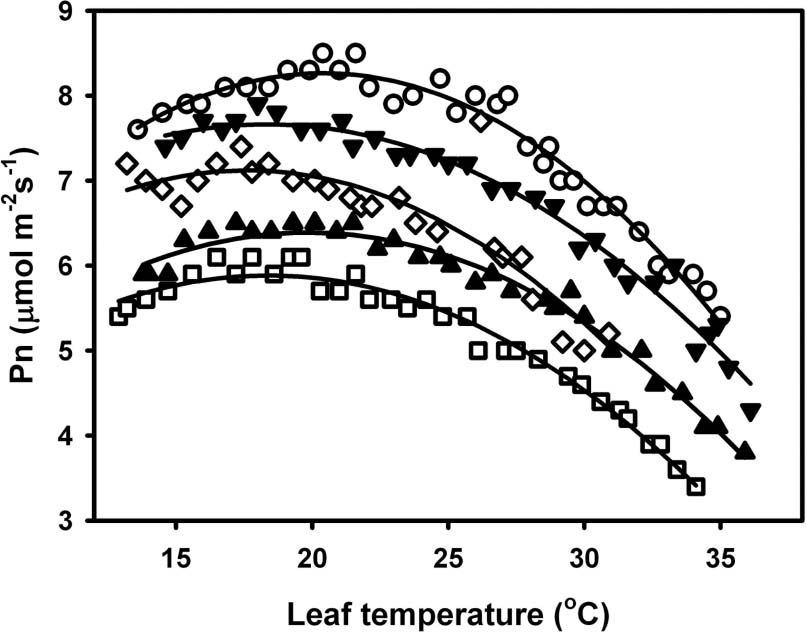
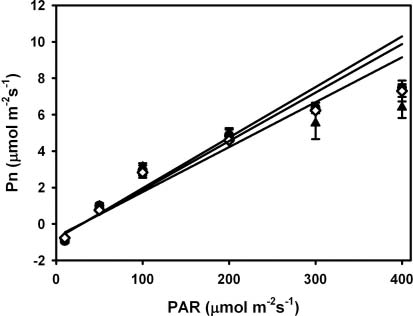
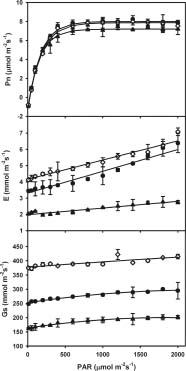
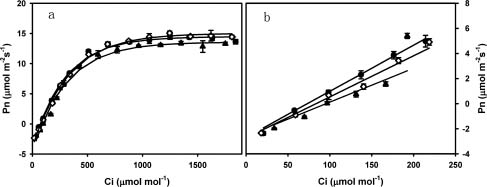
Abstract. To explore the conservation and cultivation of Cypripedium flavum at comparatively lower altitudes, a study on variations in photosynthetic characteristics was conducted on C. flavum plants from the high mountains of the eastern Himalayas. Plants of same size (6 leaves, 35-40 cm height) were selected from Tianshengqiao (alt. 3,450 m) and planted at the gardens at Zhongdian (alt. 3,200 m) and Kunming (alt. 1,900 m). The leaves of C. flavum at Tianshengqiao and Zhongdian exhibited higher photosynthetic rate, stomatal conductance, transpiration rate, apparent quantum yield, and carboxylation efficiency than their counterparts at Kunming. The optimal temperatures for photosynthesis of C. flavum plants growing at Tianshengqiao, Zhongdian, and Kunming were 18, 19 and 21°C, respectively. The optimal temperature also increased with increasing photosynthetically active radiation (PAR). However, the photosynthetic responses of C. flavum to PAR were similar at the three sites. The photosynthesis reached light saturation at the PAR of 746-832 µmol m-2s-1. The inability of C. flavum to acclimate to higher temperature resulted in the depression of photosynthetic rate at the lower altitude (Kunming). The results suggest that C. flavum adapts poorly to the warmer temperatures at low altitudes. It can be cultivated at a lower attitude, but proper agronomical methodology will need to be developed for better growth.
Keywords: Cultivation; Cypripedium flavum; Gas exchange; Optimal photosynthetic temperature; Photosynthetic capacity.
Introduction
The genus Cypripedium contains 49 species distributed in North America, East Asia, and Europe. The Hengduan Mountains are one of the most important distribution regions (Chen, 1985). Cypripedium flavum is a famous and rare alpine lady's slipper orchid that occurs in alpine grasslands or under the scrub and forest at altitudes of 2,700-3,700 m in the Hengduan Mountains (Fang, 1993). Because large-scale cultivation of this plant under artificial conditions is not economically feasible, wild populations of this species are the main source of material for horticulture. In recent years, ecological disturbance, unscientific and uncontrolled collection, tourism and increasing grazing pressure have resulted in considerable decline in the population number of C. flavum in the Hengduan Mountains (Cribb and Sandison, 1998; Huang and Hu, 2001). In order to meet the increasing demand for this species as an ornamental plant, commercial cultivation is necessary.
Strategies are being developed for the conservation and cultivation of C. flavum at lower altitudes. However, after transplanting from natural habitats to lower altitudes, C. flavum grows poorly (Weng et al., 2002), and the flower size shrinks. Previous studies showed that chilling treatment (4°C for 50 days) promotes the growth and flower
ing of Cypripedium (Olver, 1981; Kim et al., 1996), but flowering of Cypripedium depends on plant size and vegetative growth (Primack and Stacy, 1998). In fact, the growth of plant often exhibits an optimum temperature. After transplanting to lower altitude, elevated temperature decreases the photosynthetic activity by impairing physiological processes. The reduction of photosynthetic carbon fixation directly affects vegetative growth and indirectly affects flowering through decreasing plant size and the supply of nutrition (Battaglia et al., 1996; McDonald and Paulsen, 1997; Totland, 1999; Prasad et al., 2000; Iersel, 2003; Griffin et al., 2004). Although C. flavum has been studied taxonomically (Lang, 1990; Cribb, 1997; Chen et al., 1998), studies of its physiology and cultivation that would assist in domestication are lacking (Kull, 1999). Information on physiological responses of this species to changed environments, such as growth at different altitudes, is needed for applied research aiming to increase the range of cultivation of alpine ornamental plants. The ability of species to acclimate and adapt to different environments is directly/indirectly associated with their ability to acclimate at the level of photosynthesis (Pearcy, 1977), which in turn affects biochemical and physiological processes of the leaf and, consequently, the physiology and growth of the whole plant (Chandra, 2003). Our aim, in this study, was to present basic information on the photosynthesis of C. flavum in its current habitat and to compare photosynthetic rates and factors affecting it at lower altitudes. We expect that this will assist in cultivation of this species at altitudes below its natural range.
*Corresponding author. E-mail: huhong@mail.kib.ac.cn; Tel: 86-871-522 3002; Fax: 86-871-522 3005.