Bot. Bull. Acad. Sin. (2005) 46: 315-324
CHEN et al. Duplex telomeric DNA-binding proteins in Arabidopsis
Functional redundancy of the duplex telomeric DNA-binding proteins in Arabidopsis
Chung-Mong CHEN1,*, Chi-Ting WANG1, Yu-Hsin KAO1, Geen-Dong CHANG2, Chia-Hsing HO2, Feng-Ming LEE3, and Ming-Jhy HSEU4
1Institute of Plant and Microbial Biology, Academia Sinica, Taipei, Taiwan 115, Republic of China
2Institute of Biochemical Sciences, National Taiwan University, Taipei, Taiwan 106, Republic of China
3Institute of Plant Biology, National Taiwan University, Taipei, Taiwan 106, Republic of China
4Institute of Biological Chemistry, Academia Sinica, Taipei, Taiwan 115, Republic of China
(Received May 23, 2005; Accepted July 22, 2005)
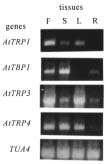
Abstract. AtTRP1 is an Arabidopsis protein that binds duplex telomeric DNA in vitro. Here we showed that knockout of AtTRP1 did not change significantly the telomere length in plant. This implies that either AtTRP1 does not participate in the regulation of telomere length or the Arabidopsis genome contains other genes functionally redundant to AtTRP1. Sequence analysis of Arabidopsis genome together with molecular cloning enabled us to identify two additional genes AtTRP3 and AtTRP4 and the corresponding cDNA clones encoding AtTRP1-like proteins. The C-terminal regions of both AtTRP3 and AtTRP4 proteins bind specifically duplex telomeric DNA in vitro. The amino acid sequence of AtTRP4 is identical to that of another Arabidopsis protein TRFL1 except with an internal deletion of six amino acids, suggesting that AtTRP4 and TRFL1 may be derived from the same gene by alternative splicing. This speculation was further confirmed by DNA sequence analysis of RT-PCR products specific for AtTRP4 and TRFL1 transcripts. Our data together with reports from other researchers revealed that Arabidopsis contains at least seven different duplex telomeric DNA-binding proteins encoded by a six-member gene family, named AtTRP. We proposed that some members of the AtTRP family may be functionally redundant in the regulation of telomere length in Arabidopsis.
Keywords: Alternative splicing; Arabidopsis thaliana; Functional redundancy; Knockout mutant; Telomere length; Telomeric DNA-binding protein.
Introduction
Telomeres are unique structures that are found at the ends of chromosomes in most eukaryotes and essential for the maintenance of the integrity of those chromosomes and for genome stability (Blackburn, 2001). Telomeric DNA consists of short DNA repeats, which are tandem arrayed and terminated with a single-stranded 3´ G-rich overhang. The synthesis of telomeric DNA at the chromosome end is primarily catalyzed by the telomerase. However, the access to telomerase is regulated by various factors, including the duplex telomeric DNA-binding proteins such as Rap1p in budding yeast, Taz1p in fission yeast, and TRF1 and TRF2 in human cells (Smogorzewska and de Lange, 2004; Vega et al., 2003).
The protein Rap1p negatively regulates telomere length (Marcand et al., 1997) in addition to controlling the transcription of multiple genes in budding yeast (Shore, 1994).
The C-terminal protein-interaction domain of Rap1p is required for the regulation of telomere length (Kyrion et al., 1992; Marcand et al., 1997) and for telomere clustering (Levy and Blackburn, 2004). It has been proposed that the recruitment of other proteins to telomere by the C-terminal domain of Rap1p can prompt the telomere to form a high-order structure inaccessible to telomerase (Levy and Blackburn, 2004).
Taz1p contains a Myb DNA-binding domain at its C-terminus and deletion of this C-terminus results in telomere lengthening (Cooper et al., 1997), indicating that Taz1p plays a negative role in the maintenance of telomere length in fission yeast. Although Taz1p was predicted to have a dimerization domain at N-terminus (Fairall et al., 2001), gel filtration analysis of purified Taz1p revealed that the native form of Taz1p can be as big as hexamers (Tomaska et al., 2004). Incubation of artificial telomeres with purified Taz1p prompted telomeric DNA to form a t-loop structure on which the associated Taz1p particles were also estimated to be hexamers (Tomaska et al., 2004). This observation implied that binding of Taz1p to telomeric DNA as oligomers may be important for t-loop formation and the regulation of telomere length in fission yeast.
*Corresponding author. Email: chen922@gate.sinica.edu.tw; Tel: (+886)-2-27899590 ext. 325; Fax: (+886)-2-27827954.
GenBank accession no.: AY181997 and AY395985.