Bot. Bull. Acad. Sin. (2005) 46: 325-331
YAN et al. Regulation of camptothecin and 10-hydroxycamptothecin levels
Tissue-specific and developmental regulation of camptothecin and 10-hydroxycamptothecin levels in Camptotheca acuminata
Xiu-Feng YAN1,*, Yang WANG1, Yu-Hong ZHANG2, Tao YU1, Mei-Fang MA1, Shi-Jie JU1, and Sixue CHEN3
1College of Life Sciences, Northeast Forestry University, Harbin 150040, P. R. China
2Open Research Laboratory of Forest Plant Ecology, Northeast Forestry University, Harbin 150040, P. R. China
3Donald Danforth Plant Science Center, St. Louis, MO 63146, U.S.A.
(Received March 26, 2004; Accepted June 24, 2005)
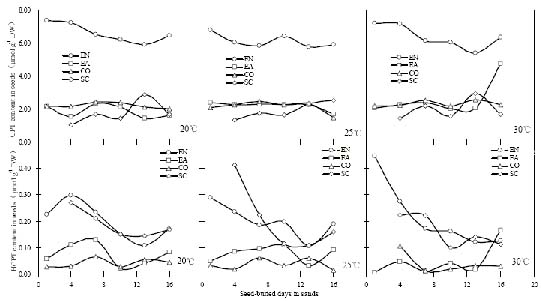
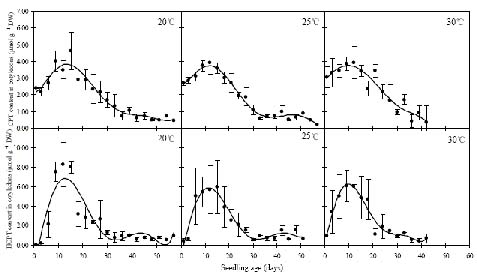
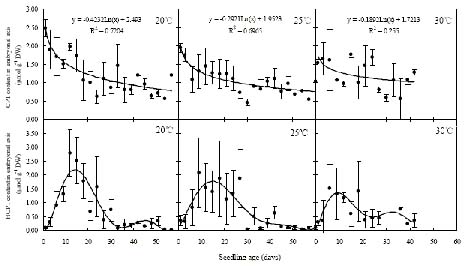
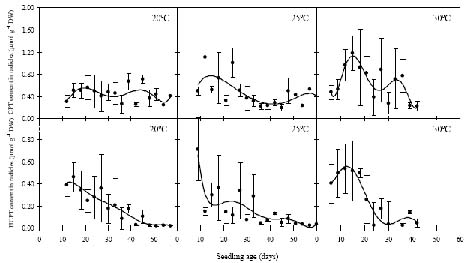
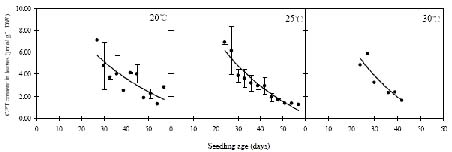
Abstract. Camptothecin (CPT) and its analogue 10-hydroxycamptothecin (HCPT) are two naturally occurring monoterpene indole alkaloids in Camptotheca acuminata. They both show potential efficacy as anti-cancer compounds as well as act as defensive chemicals in plants. Here we report the regulation of CPT and HCPT contents in C. acuminata during seed imbibition and seedling development. The levels of CPT in endosperm was 2 to 3 fold higher than in other tissues, and clearly decreased during the imbibition period. In other tissues, CPT contents exhibited slight fluctuations. In comparison, HCPT contents in all the tissues were much lower. HCPT was less abundant in cotyledon and embryonal axis than in endosperm and seed coat. During seedling development, CPT and HCPT contents in cotyledons reached maximum at 9-15 days stage. In embryonal axis, CPT contents decreased logarithmically, while HCPT content followed the same pattern as in cotyledons. CPT and HCPT levels in radicles were very low, with CPT levels remaining constant and HCPT levels decreasing during development. In leaves, the contents of CPT showed linear reduction during seedling growth. These results demonstrate that the levels of CPT and HCPT are tissue-specifically and developmentally regulated. The physiological implications and significance of the regulation are discussed.
Keywords: Camptotheca acuminata; Camptothecin; 10-hydroxycamptothecin; Regulation.
Introduction
Plant secondary metabolites are naturally occurring compounds which play important roles in various biological functions. For example, certain groups of plant secondary metabolites act as deterrents against pathogens and herbivores or attract pollinators via flower color or scent. A good example of this are alkaloids, which are a diverse group of low-molecular-weight, nitrogen- containing compounds that traditionally have been of interest due to their pronounced and diverse physiological activities in animals (Facchini, 2001). Many alkaloids are known to be poisonous to mammals and have demonstrated functions in plant defense against herbivores and pathogens (Wink, 1998).
Camptothecin (CPT), a monoterpene indole alkaloid produced by a native Chinese tree Camptotheca acuminata Decne., was isolated by Wall and co-workers (Wall et al., 1966) 39 years ago. Because of its high anti-tumor activity in the L1210 mouse leukemia assay, CPT attracted immediate attention for use as a potential cancer chemotherapeutic agent (Geran et al., 1972). The development of CPT as an anti-tumor drug stalled, due to a variety of unacceptable side effects on humans (Giovanella et al., 1989), until
inhibitor of topoisomerase I was discovered by Liu and co-workers (Hsiang et al., 1985) in 1985. Among CPT's many analogues, which have been investigated over more than 30 years, irintecan (Sawada et al., 1991; Masuda et al., 1992; Abigerges et al., 1995; Bleiberg, 1999) and topotecan (Kingsbury et al., 1991; Lilenbaum et al., 1995; Romanelli et al., 1998; Clements et al., 1999) have been approved by United States Food and Drug Administration (FDA) for application to colon/rectum and ovarian cancer. Two other related drugs, 9-aminocaptothecin and 9-nitrocamptothecin (Wani et al., 1986), are currently undergoing extensive clinical trials (Giovanella, 1997; Jeha et al., 1998; Stevenson et al., 1999). 10-hydroxycamptothecin (HCPT), a natural derivative of CPT in C. acuminata and the precursor of irinotecan and topotecan, has shown efficacy against lung, breast, and uterine cervical cancer (Zhang et al., 1998) through the same mechanism of inhibition of topoisomerase I.
Despite the importance of CPT and HCPT in medical applications, research has been conducted to study their distribution in C. acuminata (Lopez-Meyer et al., 1994; Liu and Adams, 1996; Yan et al., 2003), the induction of biosynthesis by environmental factors such as light and water (Liu et al., 1997, 1998), and the effect of seed source variations (Liu and Adams, 1998). However, little work has been done analyzing CPT and HCPT levels in different tissues during the seed imbibition or seedling development
*Corresponding author. E-mail: xfyan@mail.hl.cn; Fax: +86-451-82190052; Tel: +86-451-82192185.