Bot. Bull. Acad. Sin. (2005) 46: 333-338
NAYYAR et al. Polyamines and water stress sensitivity in Chickpea, Soybean
Involvement of polyamines in the contrasting sensitivity of chickpea (Cicer arietinum L.) and soybean (Glycine max (L.) Merrill.) to water deficit stress
Harsh NAYYAR*, Satwinder KAUR, SMITA, Sanjeev KUMAR, K.J. SINGH, and K.K. DHIR
Department of Botany, Panjab University, Chandigarh 160 014, India
(Received December 29, 2004; Accepted May 13, 2005)
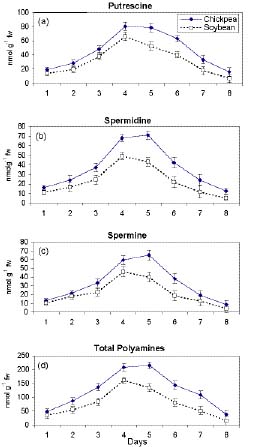
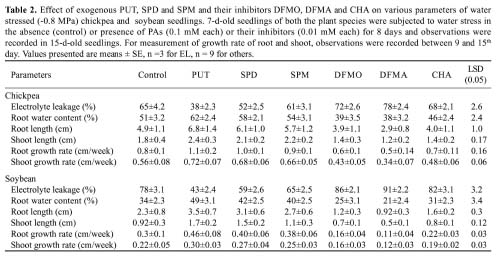
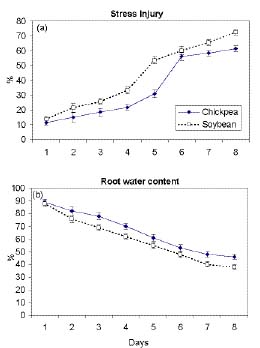
Abstract. Chickpea (Cicer arietinum L. cv. GPF2) and soybean (Glycine max (L.) Merrill cv. Brag) genotypes were subjected to varying water stress levels of -0.2 to -0.8 MPa to assess their relative sensitivity towards water stress. Soybean seedlings experienced significantly more stress injury (as electrolyte leakage) than chickpea at all stress levels. LD50 in terms of percent germination, root length, and root growth rate occurred at -0.4 MPa in soybean and at -0.6 MPa in chickpea. In chickpea, the root water content was higher than in soybean during stress. Endogenous levels of individual as well as total polyamines (PAs) in roots of 7-d-old seedlings subjected to -0.8 MPa stress increased to a significantly greater extent in chickpea than in soybean. The stress injury was accentuated as the PA levels declined in both the plant types. The reduced levels of PAs in soybean, especially putrescine (PUT) and spermidine (SPD) relative to chickpea, were related to higher stress injury and decreased water content. Exogenous PUT and SPD markedly mitigated the stress-induced effects, particularly in soybean. Inhibitor studies involving a-Difluromethylarginine (DFMA) and a-difluromethylornithine (DFMO), the biosynthetic inhibitors of PUT, as well as cyclohexylamine (CHA), biosynthetic inhibitor of SPD and SPM, corroborated the role of PAs in mediating the differential sensitivity of chickpea and soybean to water stress.
Keywords: Cicer arietinum; Glycine max; Polyamines; Putrescine; Spermidine; Spermine; Water stress.
Abbreviations: CHA, Cyclohexylamine; DFMA, a-Difluromethylarginine; DFMO, a-difluromethylornithine; EL, Electrolyte leakage; PAs, Polyamines; PUT, Putrescine; SPD, Spermidine; SPM, Spermine.
Introduction
Chickpea (Cicer arietinum L.) and soybean (Glycine max (L.) Merrill), the two most important pulses of India are grown as rainfed crops in the country, and consequently they experience water deficit stress at one or the other growth stage(s). A distinctive variation exists between the two plant types in their sensitivity to water stress (Grzesiak et al., 1996). Chickpea is considered relatively more tolerant, possibly because its root system is deeper, its leaves and canopy smaller than soybean (Serraj et al., 2004). The metabolic reasons governing their differential sensitivity towards water stress are not known. Moreover, the contrasting responses of different plant types to water stress offer an excellent model to elucidate the underlying causes of stress sensitivity, which can be employed to develop better stress tolerance in sensitive crops.
Several adaptive mechanisms are evoked by plants in response to water stress (Chaves et al., 2003). Interest has been growing in the possible involvement of polyamines (PAs) in the defense reaction of plants to various environmental stresses (Kao, 1997; Bouchereau et al., 1999;
Kakkar and Sawhney, 2003). PAs are polycationic cellular molecules that play an essential role in cell growth and differentiation (Evans and Malmberg, 1989), and at a physiological pH, PAs can bind strongly to the negative charges in cellular components such as nucleic acids, proteins, and phospholipids (Smith, 1985). Interactions of PAs with membrane phospholipids may stabilize the membranes under conditions of stress (Roberts et al., 1986). It has been found that stress-tolerant plants increase their endogenous PAs levels to a much greater extent than sensitive ones (Lee, 1997). Furthermore, transgenic plants overproducing PAs possess greater stress tolerance (Galston et al., 1997), and exogenous PAs confer protection from a variety of abiotic stresses (Basra et al., 1997; Nayyar and Chander, 2004). In the present study, we tested the hypothesis that the variation in water stress sensitivity between chickpea and soybean plants is related to changes in the levels and functioning of endogenous PAs.
Materials and Methods
Plant Material and Growth Conditions
Seeds of chickpea (Cicer arietinum L. cv. GPF2) and soybean (Glycine max (L.) Merrill cv. Brag) procured from Panjab Agricultural University, Ludhiana, India, were surface sterilized with 0.1% mercuric chloride for 2 min and subsequently washed thoroughly with distilled water.
*Corresponding author. E-mail: nayarbot@pu.ac.in; harshnayyar@hotmail.com