Botanical Studies (2010) 51: 521-529.

Alteration of phytoplankton assemblages caused by changes in water hardness in Feitsui Reservoir, Taiwan
Jiunn-Tzong WU1,2'* and Lai-Chu KOW1
1Biodiversity Research Center, Academia Sinica, Taipei 11529, Taiwan
2Institute of Ecology and Evolutionary Biology, National Taiwan University, Taipei 10660, Taiwan
2Institute of Ecology and Evolutionary Biology, National Taiwan University, Taipei 10660, Taiwan
(Received March 8, 2010; Accepted May 14, 2010)
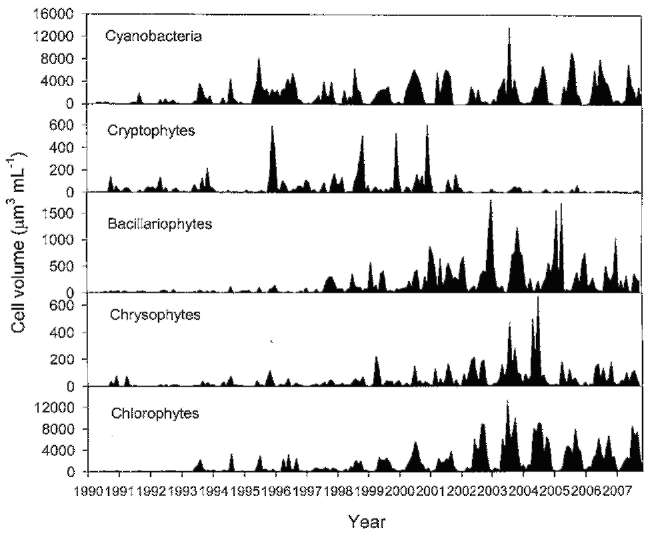
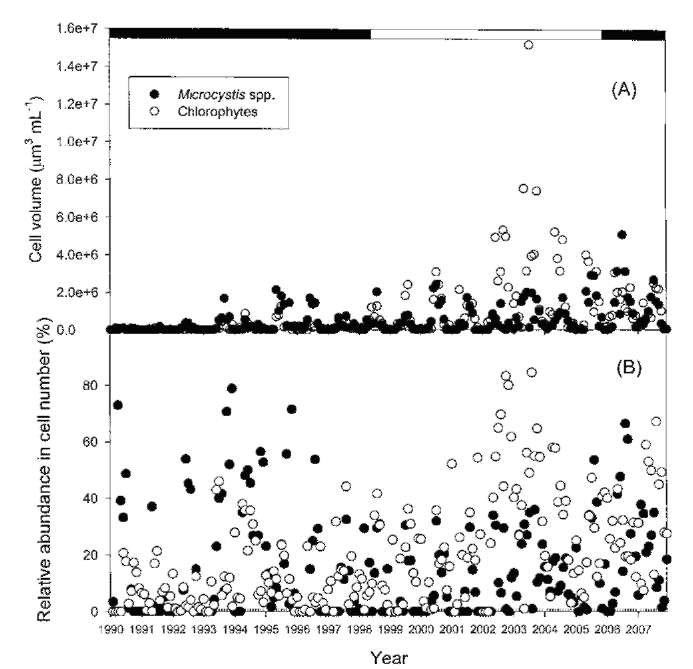
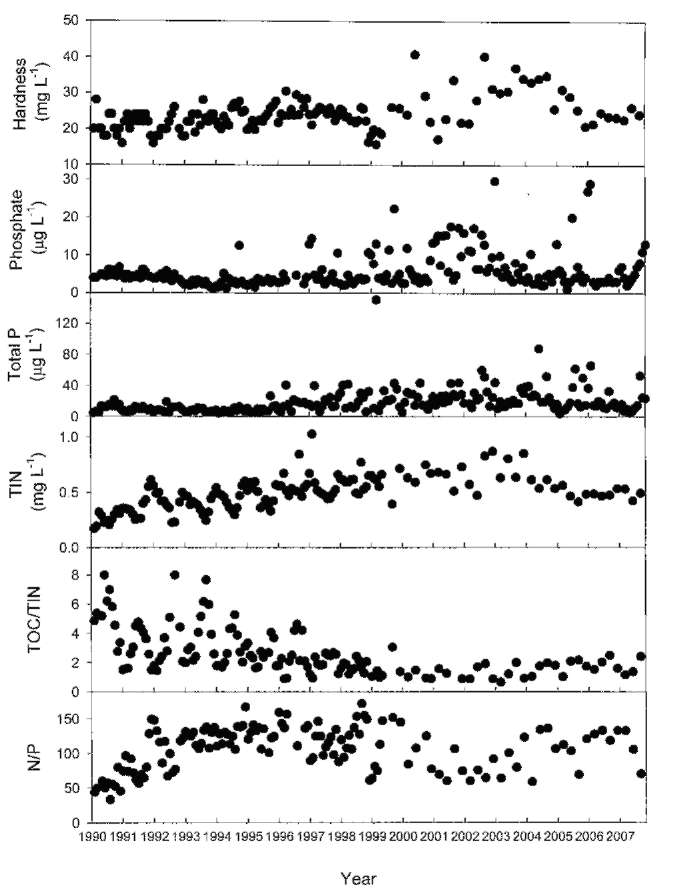
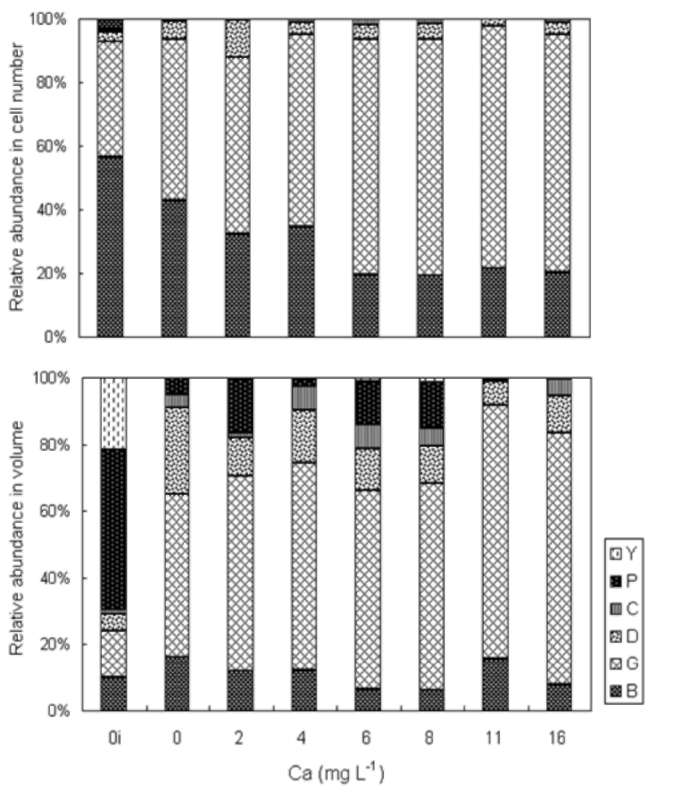
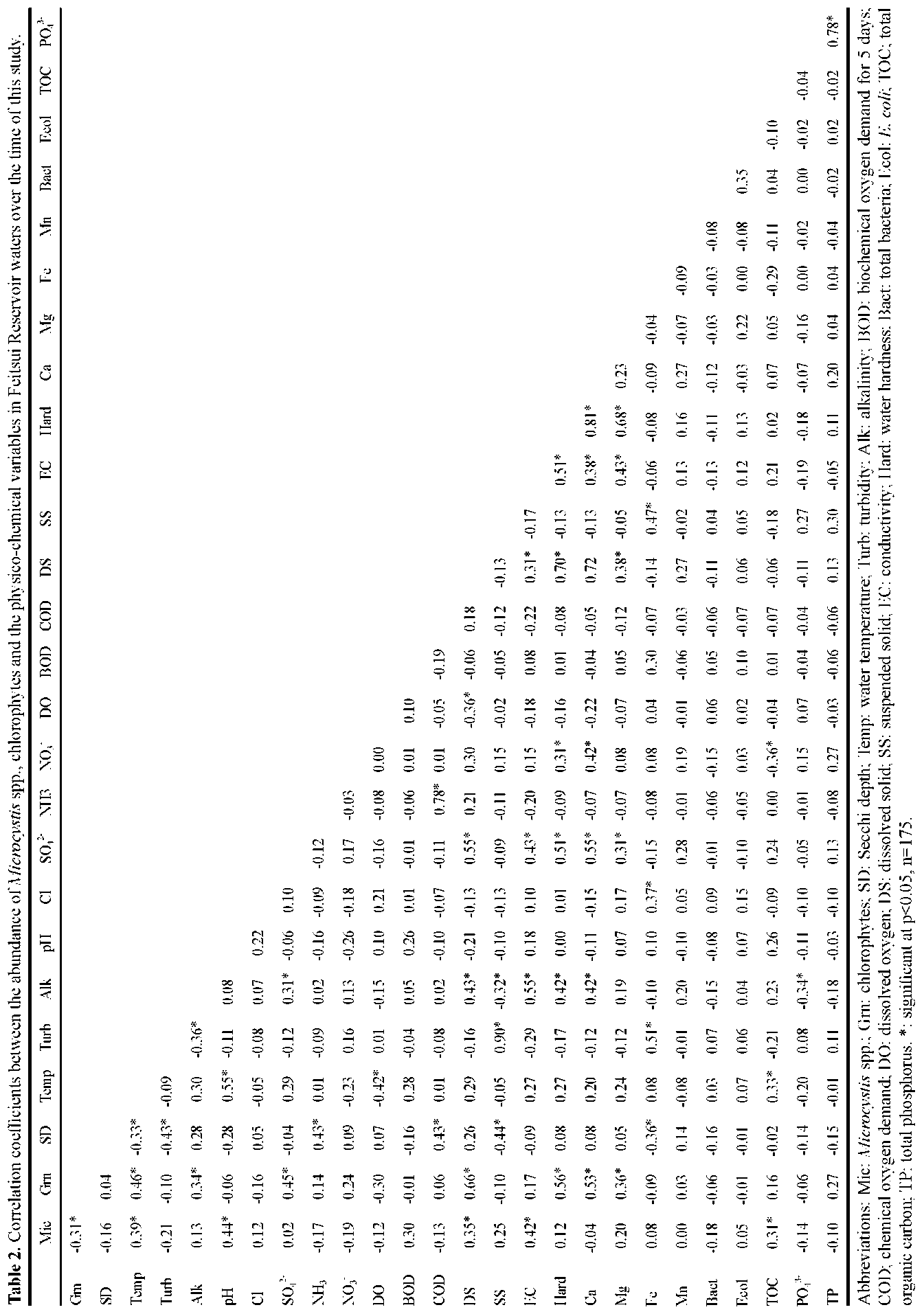
ABSTRACT. Feitsui Reservoir is a subtropical water reservoir constructed in 1986 for supplying the domestic water of Taipei metropolitan area. During long-term monitoring conducted from 1990 to 2007, we observed a remarkable alteration in dominant phytoplankton groups, showing a succession of chlorophytes versus Microcystis spp. Analysis of such change with respect to 27 physico-chemical variables in reservoir waters showed that the either phytoplankton group displayed different nutrient demands and that the dominance of chlorophytes was closely related to the elevated level of water hardness, including calcium, magnesium, soluble solids, and sulfate, which were resulted from the pollutants discharged from the construction site of highway in the watershed. A calcium-enrichment experiment supported the belief that the higher chlorophyte density was due to elevated calcium levels. The possible way by which calcium influenced the availability of carbon- as well as phosphate-source in waters was discussed. This study suggests the possible strategy of mitigating the Microcystis problems in water reservoir.
Keywords: Ca-enrichment; Feitsui Reservoir; Phytoplankton; Subtropical water reservoir; Water hardness.
INTRODUCTION
with the occurrence of Microcystis in FTR.
Over last two decades, two important events occurred in the watershed that disturbed the aquatic environment of FTR, the construction of a wastewater treatment plant in 1994 to treat the sewage waters discharged from the villages and the construction of a highway across the watershed, which began in 1996 and was completed in 2005. Both construction sites were very close (<2 km) to the major riverine inflow to the FTR. The turbid wastewater discharged from the construction sites contained clay (suspended solids), soluble solids, and a variety of nutrients including phosphorus, ammonia, and cations including calcium and magnesium (Wu and Kow, 2002). The concentrations of these compounds were particularly high in the wastewater discharged in the area in which a 12.9 km tunnel was constructed. Even though a portion of the wastewater was treated prior to being discharged, the majority of the pollutants entered the FTR. As a result, these construction projects have had a large impact on the water quality as well as the structure of the phytoplankton community in the reservoir. In this study, we attempted to identify key factors that regulate these changes in the phytoplankton community. To accomplish this, we evaluated data pertaining to 27 physio-chemical variables that were collected from 1990 to 2007 on a monthly basis. In addition, we conducted a calcium-enrichment experiment to confirm the factors identified by the data analysis.
In Taiwan, an increase in the input of anthropogenic pollutants to the aquatic environment has resulted in eutrophication of a number of water resources. As a result, problems associated with eutrophication such as the presence of toxins and off-flavors in drinking water have become a common occurrence. Such nuisance problems are usually related to the presence of certain harmful cyanobactaria (Wu and Hsu, 1988; Carmichael, 2001; Huisman et al., 2005). As a result, the control of cyanobacteria has received great attention and interest in recent years (Chorus and Bartram, 1999; Kim, 2008).
Feitsui Reservoir (FTR) is a water reservoir which was constructed in 1986 in order to supply domestic water to the Taipei metropolitan area. Long-term monitoring of the water quality and phytoplankton in this reservoir has been conducted since its construction (Wu and Kow, 2008). During the monitoring period, a Microcystis bloom occurred in 1988. Since then, high concentrations of this cyanobacterium have existed in the FTR. M. aeruginosa is one of species which has been shown to produce the hepatotoxin microcystin-LR (Liu, 1995). As a result, a great attention has been paid to the problems associated
*Corresponding author: E-mail: jtwu@gate.sinica.edu.tw; Tel:+886-2-27871073; Fax: +886-2-27871182.