|
|
|
|
|
|
|
|
|
|
|
|
Botanical Studies (2011) 52: 105-113
|
|
|
|
|
|
|
|
|
Sinosenecio nanchuanicus (Asteraceae), a new species small in size yet high in chromosome number from Chongqing, China
|
|
|
|
|
|
|
|
Zheng-Yu LIU2, Ying LIU3, and Qin-Er YANG1*
|
|
|
|
|
|
|
|
1Key Laboratory of Plant Resources Conservation and Sustainable Utilization, South China Botanical Garden, Chinese
Academy of Sciences, Xingke Road, Tianhe District, Guangzhou 510650, China
2Chongqing Institute of Medicinal Plant Cultivation, Sanquan, Nanchuan, Chongqing 408413, China
3State Key Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, Beijing
100093, China
|
|
|
|
|
|
|
|
(Received September 4, 2009; Accepted August 20, 2010)
|
|
|
|
|
|
|
|
ABSTRACT. Sinosenecio nanchuanicus Z. Y. Liu, Y. Liu & Q. E. Yang, a new species of the Asteraceae from Chongqing, China, is described and illustrated. Its chromosome number (2n = 144) is reported here. This new species is similar to S. bodinieri (Vaniot) B. Nord., a diploid species with 2n = 48, in the posture, the ro-tundate or subrotundate leaf-lamina, the glabrous ovaries, and the pappose achenes, but differs in the smaller stature, the smaller leaf-lamina shallowly regularly 5-7-lobed and dark green above, the veins sunken above, the capitula constantly solitary and terminal, the rays 14-15, and the high chromosome number as well. From S. palmatilobus (Kitam.) C. Jeffrey & Y.L. Chen it differs also by these characters and, additionally, by its leaf-lamina rotundate or subrotundate in outline (vs. broadly ovate) and short-ciliate along the margin (vs. long-ciliate). All the taxa put in synonymies under S. bodinieri and S. palmatilobus were carefully re-examined and compared to S. nanchuanicus. Line drawings, distribution map, light microscope (LM) microphotographs of floral characters of S. nanchuanicus, color photographs of S. bodinieri, S. nanchuanicus and S. palmatilobus are provided.
|
|
|
|
|
|
|
|
Keywords: Asteraceae; Chromosome number; Limestone flora; Local endemism; Polyploidy; Senecioneae; Sinosenecio nanchuanicus.
|
|
|
|
|
|
|
|
|
regularly 5-7-lobed leaf-lamina, the veins sunken above, and the constantly solitary, terminal capitula. Field observations carried out in the Jinfoshan Mountain area and some other sites confirmed and increased the list of differences between the plant and S. bodinieri, leading us to recognize it as a new species. Further herbarium work resulted in the discovery of one more herbarium specimen, Zheng-yu Liu 771310 (Figure 1A), which is designated as the holotype of the new species. The results of our cyto-logical studies indicate that the new species had an unusually high chromosome number (2n = 144) in the genus, lending strong support to our taxonomic treatment.
|
|
|
|
For the second author's Ph.D. project on the systemat-ics and evolution of Sinosenecio B. Nord. (Senecioneae, Asteraceae), we made a comprehensive survey of the specimens of this genus deposited in Chinese herbaria. One fruiting collection kept in the Herbarium of Institute of Botany, Chinese Academy of Sciences (PE), You-sheng Chen 6142, made from the Jinfoshan Mountain, Nanchuan County, Chongqing Municipality, China, caught our attention. At first glance, this plant, because of its small size, appeared to be only an extreme variant of S. bodinieri (Vaniot) B. Nord., a quite polymorphic species widely distributed in China and very common in the Jinfoshan Mountain area, owing to similarities in the posture, the rotundate or subrotundate leaf-lamina and the pappose achenes. Upon a closer examination, however, we found that the plant in question differs from S. bodinieri in the
|
|
|
|
|
|
|
|
Sinosenecio nanchuanicus Z. Y Liu, Y. Liu & Q. E. Yang, sp. nov.―TYPE: CHINA. Chongqing, Nanchuan County, Jinfoshan Mountain, Da-fo-yan (literally meaning "Big Buddha Rock"), alt. 1,250 m, on rock cliff along margin of evergreen broad-leaved forests, 26 May 1977, Zheng-yu Liu 771310 (holotype, IBSC).南川蒲兒根
Figures 1, 2, 3
|
|
|
|
*Corresponding author: E-mail: qeyang@scib.ac.cn; Tel: 8620-37094273; Fax: 86-20-37094273.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Botanical Studies, Vol. 52, 2011
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Figure 1. Sinosenecio nanchuanicus. A, China, Chongqing, Nanchuan, Jinfoshan, Zheng-yu Liu 771310 (holotype, IBSC); B, China, Chongqing, Nanchuan, Jinfoshan, Ying Liu & Tao Deng 2008035 (paratype, IBSC).
|
|
|
|
|
|
|
|
|
Ob habitum, foliorum laminas rotundatas vel subro-tundatas, ovaria glabra et achenia pappo instructa haec species arcte similis Sinosenecioni bodinieri (Vaniot) B. Nord., sedplanta breviore, 10-12 cm alta, foliorum laminis regulariter 5-7-lobatis, minoribus, 0.8-1.7 x 0.8-2 cm, supra atro-viridibus, venis supra impressis, capitulis singularibus terminalibus, ligulis 14-15, numero chromo-somatum (2n = 144) distinguitur.
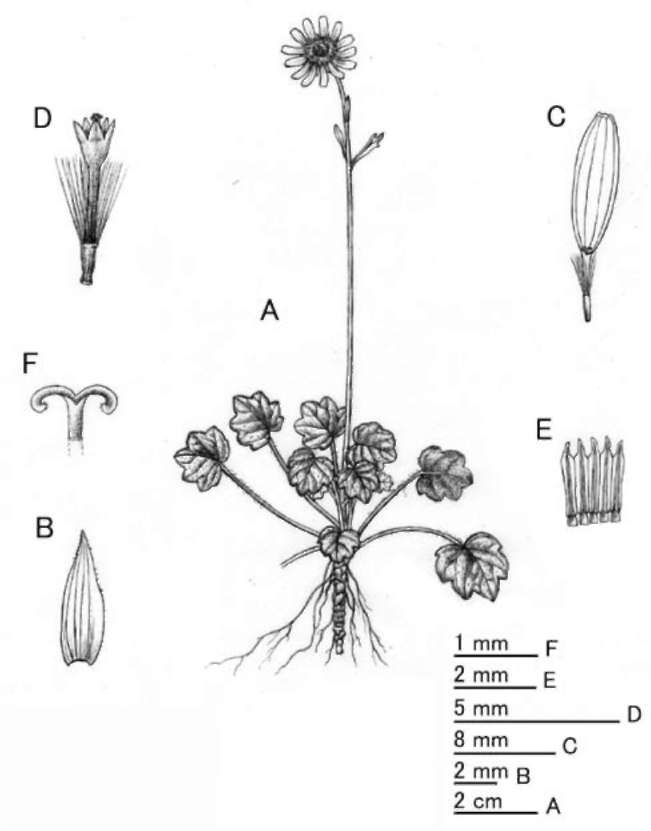
Scapigerous herb; stolons absent. Rhizomes 3-4 mm in diameter. Stems solitary, erect, 10-12 cm tall, simple, purple, glabrescent, bearing several small spathulate leaf-like bracts in upper part. Leaves several, radical, long-petiolate; lamina rotundate or subrotundate in outline, apex acute, base shallowly cordate, margin short-ciliate, 0.8-1.7 x 0.8-2 cm, shallowly regularly 5-7-lobed with broadly deltoid or rounded, entire, apiculate lobules, subcoriaceous, nitid, dark-green above, purple beneath, pubescent above, sparsely pubescent or glabrescent on the midvein beneath, palmately veined, veins sunken above; petioles 1-4 cm long, pubescent, base expanded, not auriculate. Capitula solitary, terminal, peduncles sparsely pubescent or glabres-cent in upper part. Involucres campanulate, ecalyculate, 7 x 9 mm. Phyllaries 13, uniseriate, oblong-lanceolate, ca. 6 x 1.5 mm, apex acuminate or acute, sparsely pubescent, fimbriate-ciliate at the apex, herbaceous, purple in upper part, green at the base. Ray florets 14-15; corolla tube 1.5 mm long, glabrous; rays yellow, oblong-elliptic, ca. 9 x 1.5 mm, apically 3-denticulate, 4-veined. Disc florets many;
|
|
|
|
Figure 2. Sinosenecio nanchuanicus Z. Y Liu, Y Liu & Q. E. Yang. A, Habit; B, Phyllary; C, Ray floret; D, Disc floret; E, Anthers; F, Style-arms (All from Zheng-Yu Liu 771310, IBSC).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
LIU et al. ― Sinosenecio nanchuanicus, a new species from China
|
|
|
|
|
|
|
|
|
|
three populations of S. nanchuanicus are located within the Jinfoshan Natural Reserve, the species' survival is still under threat because of its small population size and habitat disturbance via human activities such as medicinal plant cultivation and sightseeing.
Floral micromorphological characters. For
observation of the anther endothecial cell wall thickenings and filament collar of Sinosenecio nanchuanicus, heads were boiled in distilled water for 3 min, and then fixed with Carnoy I (glacial acetic acid: absolute ethanol = 1:3). Mature disc florets removed from the fixed heads were dehydrated in 70% ethanol for 30 min and then in 99% ethanol for 1 h before they were treated with 5% NaOH overnight. The anther tissue was isolated from the florets on the slide, flooded with 50% glycerol and a cover slip was applied. Samples were then examined at 200x (filament collar) and 400x (endothecial cell wall thickenings) magnification by light microscopy and photographed.
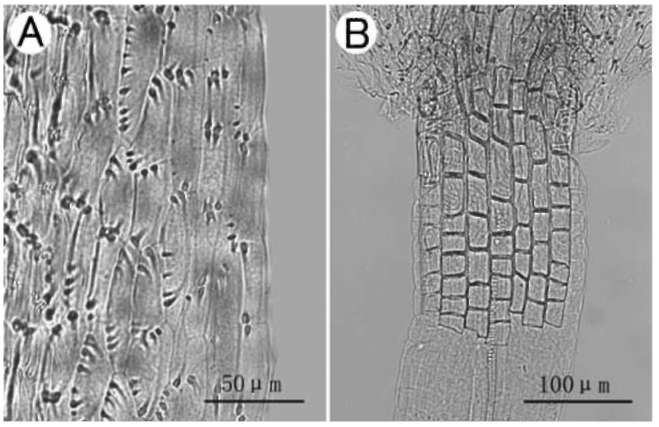
The anther endothecial cell wall thickenings of Sino-senecio nanchuanicus were polar and radial (Figure 5A), with the cells of radial thickenings being predominant,
|
|
|
|
|
|
|
|
Figure 3. Sinosenecio nanchuanicus. A, Posture; B, Capitu-lum (above) and florets (below); C, Leaf (above: adaxial side; below: abaxial side); D, Habitat (All from Jinfoshan, Nanch-uan, Chongqing, China and vouched by Ying Liu & Tao Deng 2008035, IBSC).
corolla ca. 4 mm long, tube 3 mm long, limb campanu-late; lobes ovate-lanceolate. Anthers ca.1.5 mm long, base obtuse, appendages ovate-oblong. Style arms ca. 0.5 mm long, apex truncate. Achenes cylindrical, ca. 1 mm long, smooth, glabrous. Pappus white, ca. 2.5 mm long, persistent.
Additional specimens examined. CHINA. Chongqing, Nanchuan County, Jinfoshan Mountain, Tou-ping, alt. 1,700 m, on rock cliff under waterfall, 4 May 2007, Qin-er Yang, Qiong Yuan & Ying Liu 703 (IBSC); Nanchuan County, Jinfoshan Mountain, the same locality, 4 Jun 2008, Ying Liu & Tao Deng 2008035 (IBSC); Nanchuan County, Jinfoshan Mountain, alt. 1,800 m, 25 Jul 2006, You-sheng Chen 6142 (PE).
Etymology. The specific epithet 'nanchuanicus' is derived from the type locality, Nanchuan County, Chongqing Municipality, China.
Phenology. Flowering from late May to June; fruiting June.
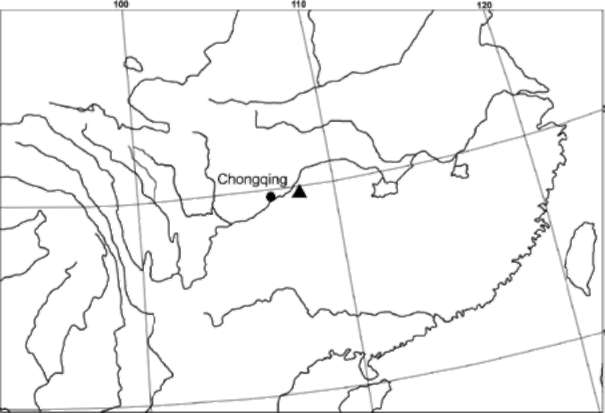
Distribution and habitat. Sinosenecio nanchuanicus is currently known from three populations in the Jinfoshan Mountain in Nanchuan County, Chongqing Municipality, China (Figure 4), growing on rock cliffs under waterfall or along margin of evergreen broad-leaved forests at altitudes between 1,200-1,800 m above the sea level. Although the
|
|
|
|
Figure 4. Distribution of Sinosgmcio nanchuanicus (▲).
|
|
|
|
|
|
|
|
Figure 5. Anther endothecial cell wall thickenings (A) and filament collar (B) of Sinosenecio nanchuanicus. A, Polar and radial thickenings; B, Uniformly sized cells (All from Jinfoshan, Nanchuan, Chongqing, China and vouched by Ying Liu & Tao Deng 2008035, IBSC).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Botanical Studies, Vol. 52, 2011
|
|
|
|
|
|
|
|
conforming to the results reported previously for some species in Sinosenecio, such as S. koreanus (Kom.) B. Nord. and S. septilobus (Chang) B. Nord. (Jeffrey and Chen, 1984). Sinosenecio bodinieri, the putative close relative of S. nanchuanicus, had also the same polar and radial pattern of endothecial cell wall thickenings (the results not shown here). As shown in Figure 5B, the filament collar of S. nanchuanicus consisted of uniformly sized cells, which is one of the diagnostic features of the genus (Nordenstam, 1978; Jeffrey and Chen, 1984; Chen, 1999).
Chromosome cytology. As we failed to harvest actively growing roots both in field and glasshouse for chromosomal observation, leaf buds were used. They were pre-treated with 0.1% colchicine for 1.5-2 h before being fixed in Carnoy I (glacial acetic acid: absolute ethanol = 1 : 3), then macerated in a 1:1 mixture of 45% acetic acid and 1 M HCl at 60°C for 3 min, stained and squashed in Carbol fuchsin.
One population (Ying Liu & Tao Deng 2008035, IBSC)
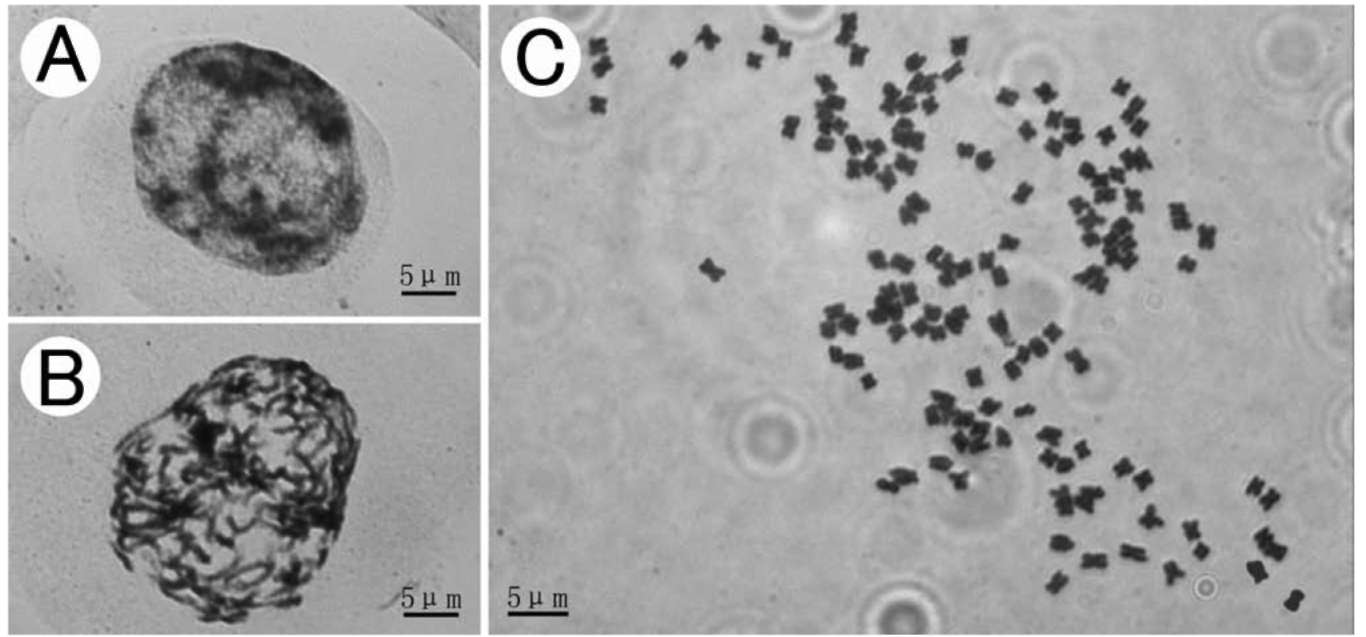
of Sinosenecio nanchuanicus was cytologically studied. In the interphase nucleus, a few darkly stained condensed bodies were observed, but their boundaries were not clear, because the other part was also stained fairly well but unevenly (Figure 6A). The prophase chromosomes displayed a distinctly continuous condensation pattern (Figure 6B). The metaphase chromosomes were counted to be 2n = 144 (Figure 6C). The chromosome number of its putative close relative, S. bodinieri, counted from material collected from Nanchuan, Chongqing, is 2n = 48 (the results not shown here). From the results of our extensive cytologi-cal studies (Ying Liu and Qin-er Yang, unpublished data), the lowest basic chromosome number for the genus previously assumed by Liu (1999), x =12, has been confirmed to be wrong, but x = 24 has been determined as one of the commonest basic numbers [the other commonest one
|
being x = 30 (Liu and Yang, 2010)]. As such, among the species of the genus with available chromosomal data, S. jishouensis D. G. Zhang, Y. Liu & Q. E. Yang, with 2n =
48 and 96 (Zhang et al., 2008), includes both diploid (2x)
and tetraploid (4x) cytotypes, S. baojingensis Y. Liu & Q. E. Yang and S. hupingshanensis Y. Liu, D. G. Zhang & Q. E. Yang, both with 2n = 48 (Liu and Yang, 2009; Liu et al., 2010), are diploids, and the new species S. nanchuanicus, with an unusually high number of 2n = 144 in the genus, is a hexaploid (6x), the highest ploidy level thus far known from the genus.
Notes. In the genus Sinosenecio, S. bodinieri is a relatively widespread and rather polymorphic species, so a somewhat detailed account of its morphological variation and taxonomy may help to gain a better understanding of the demarcation between this species and our new species S. nanchuanicus. When Vaniot (1902) described Senecio bodinieri, he pointed out that this species fell into three varieties: var. brevior, with the stem not surpassing the leaves in height and all the leaves being radical, var. elatior, with the stem obviously surpassing the leaves in height and borne with a leaf, and the radical leaves shal-lowly lobate, and var. parcepilosa, which was different from the second variety only by the plant being more sparsely pilose. Handel-Mazzetti (1937) described another variety under the species, i.e. var. elatissimus Hand.-Mazz. Lauener (1976) reduced Vaniot's three varieties just mentioned above, together with S. conferviferus H. Lev., to the synonymy of S. bodinieri, and designated a collection made from the Huangguoshu (Hoang-ko-chou) Waterfall, Guizhou, i.e. Martin & Bodinier 2572 (E) (Figure 7A), as its lectotype. Because this specimen is also the holotype of S. bodinieri var. brevior, obviously this variety was regarded by Lauener (1976) as the typical variety of S. bodinieri. In this work Lauener did not mention var. elatissimus.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Figure 6. Interphase nucleus (A), mitotic prophase (B), metaphase (C, 2n = 144) chromosomes of Sinosenecio nanchuanicus (All from Ying Liu & Tao Deng 2008035, IBSC).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
LIU et al. ― Sinosenecio nanchuanicus, a new species from China 109
|
|
|
|
|
|
|
|
Nordenstam (1978), however, when establishing Sinosene-cio, transferred Senecio bodinieri, var. brevior, var. elatior, and var. elatissimus all into this genus, raised the varieties to specific rank, and proposed four new combinations, viz. Sinosenecio bodinieri (Vaniot) B. Nord., S. brevior (Van-iot) B. Nord., S. elatior (Vaniot) B. Nord., and S. elatiss-mus (Vaniot) B. Nord. For some unknown reason, he did not mention and treat var. parcepilosa, which was regarded later by Kitamura (1982) to be worthy of a specific status, and was named Senecio palmatilobus Kitam. In their monographic revision of the genus Sinosenecio, Jeffrey and Chen (1984) essentially adopted Lauener's concept on Senecio bodinieri, reducing Sinosenecio brevior, S. elatior, S. elatissmus and Senecio conferviferus to its synonymy,
|
but they followed Kitamura (1982) in recognizing the specific status of var. parcepilosa, and proposed a new combination Sinosenecio palmatilobus (Kitam.) C. Jeffrey & Y. L. Chen. They pointed out explicitly that Senecio bodinieri var. brevior is a name not correctly published (nom. non rite public.) and that Sinosenecio brevior is a superfluous name (nom. superfl.). From the above account it is clear that the Senecio bodinieri group defined by Vaniot (1902) is currently treated as including two species, S. bodinieri and S. palmatilobus. According to Jeffrey & Chen (1984), and Chen (1999), these two species differ mainly in the shape of the leaf-lamina base, with the leaf-lamina in S. bodinieri being cordate at the base whereas that in S. pal-matilobus being truncate at the base.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Figure 7. Some specimens of the Sinosenecio bodinieri group with the leaf-lamina broadly ovate in outline and long-ciliate along the
and Sinosenecio brevior, lectotype of Senecio bodinieri and Sinosenecio bodinieri; B, China, Guizhou, Huangguoshu, Qin-er Yang, Qiong Yuan & Ying Liu 682 (IBSC), a collection showing no essential differences from Martin & Bodinier 2572 (E); C, China, Gui-zhou, near Hin-y-hien (Xingyi), Bodinier 1539 (E), holotype of Senecio bodinieri var. parcepilosa, isotype of Senecio palmatilobus and Sinosenecio palmatilobus; D, China, Guizhou, Xingyi, Qin-er Yang, Qiong Yuan & Ying Liu 673 (IBSC), a collection showing no essential differences from Martin & Bodinier 2572 (E).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Botanical Studies, Vol. 52, 2011
|
|
|
|
|
|
|
|
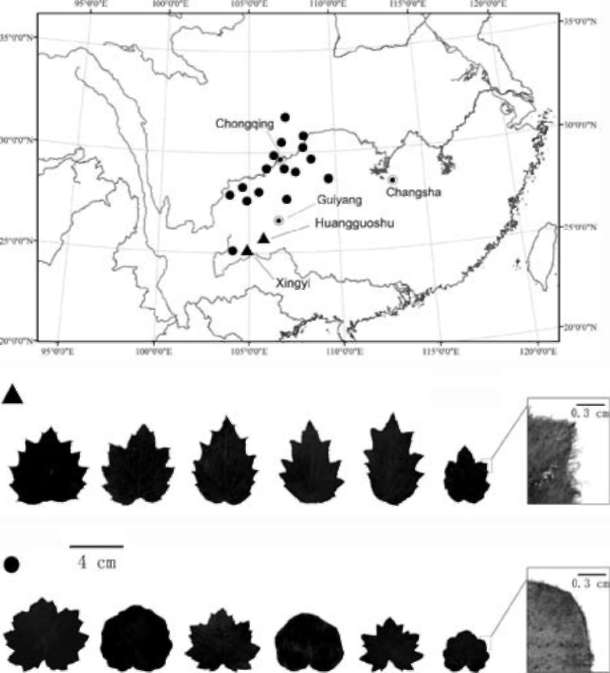
After examining a large number of relevant specimens and conducting field observations in some sites, such as Huangguoshu Waterfall, Guizhou (type locality of Senecio bodinieri var. brevior and Sinosenecio bodinieri), Jinfos-han, Chongqing (type locality of Senecio bodinieri var. elatissmus and Sinosenecio nanchuanicus), and Xingyi, Guizhou (type locality of Senecio bodinieri var. parcepi-losa and Sinoseneco palmatilobus), we agree with Lauener (1976), Jeffrey and Chen (1984), and Chen (1999) that such character as the relative length of the stem and the radical leaves is not a good taxonomic character in this group of taxa (Figures 7, 8). We also found that the shape of the leaf-lamina base may not be a reliable distinguishing character for the group, in which the lamina base exhibits remarkable variation, being truncate, shallowly cordate to deeply cordate as illustrated in Figure 9. The outline of the leaf-lamina and the length of the ciliae along the leaf-lamina margin, however, seem to be correlated with the geographical distribution and thus may be two better
|
diagnostic characters (Figure 9). To make matters even more complicated, among the populations investigated, the Huangguoshu population and the Xingyi population, both in Guizhou Province, were found to be very similar to each other in the leaf-lamina being broadly ovate in outline and long-ciliate along the margin; through these two characters they can be separated from all the other populations of the group which are characterized by the leaf-lamina being rotundate or subrotundate in outline and short-ciliate along the margin. This variation pattern indicates that Sinosenecio bodinieri and S. palmatilobus may better be treated as belonging to a same taxonomic entity which is currently known only from Huangguoshu and Xingyi in Guizhou, whereas S. elatior, S. elatissimus and Senecio conferviferus may be treated as belonging to another one, which has been found to occur in Chongqing, Guizhou, Hunan, Sichuan, and Yunnan. Clearly this change in classification will result in some changes in nomenclature, which, depending on the taxonomic rank given to the two
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Figure 8. Some specimens of the Sinosenecio bodinieri group with the leaf-lamina rotundate or subrotundate in outline and short-ciliate along the margin. A, China, Guizhou, near Kouy-yang, Bodinier 2197 (E), syntype of Senecio bodinieri var. elatior, holotype of Senecio conferviferus; B, China, Chongqing, Nanchuan, Wen-pei Fang 833 (E), holotype of Senecio bodinieri var. elatissimus; C, China, Chongqing, Nanchuan, Jinfoshan, Ying Liu & Tao Deng 2008033 (IBSC), showing two smaller plants; D, China, Chongqing, Nanchuan, Jinfoshan, Ying Liu & Tao Deng 2008031 (IBSC), showing a smaller plant and a larger one; E, China, Chongqing, Nanchuan, Jinfoshan, Qin-er Yang, Qiong Yuan & Ying Liu 699 (IBSC, PE), showing a larger plant.
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
LIU et al. ― Sinosenecio nanchuanicus, a new species from China
|
|
|
|
|
|
|
|
|
entities, would mainly involve choosing a correct name for the entities each and re-deposing some synonyms. As the authenticity of the variation pattern as elaborated above needs to be further confirmed by more studies, and a reclassification of this group of taxa is beyond the scope and intention of the present study, we here do not formally re-classify S. bodinieri and S. palmatilobus but provisionally follow Jeffery and Chen's (1984) treatment. It seems that these two species are much more closely related to each other than previously regarded and not always readily distinguishable. Anyhow, our new species S. nanchuanicus does not match well with any variant forms in the S. bodi-nieri group. From all these variants S. nanchuanicus can be separated by a character combination: the often smaller stature, the smaller leaf-lamina shallowly regularly 5-7-lobed and dark green above, the veins sunken above, the capitula constantly solitary and terminal, and the rays 1415. Therefore, no matter what taxonomic changes may be made on the S. bodinieri group in future, the validity of S. nanchuanicus seems not to be affected.
In Table 1, Sinosenecio nanchuanicus is compared with S. bodinieri [sensu Jeffrey and Chen (1984)] in detail. Sinosenecio nanchuanicus is very similar to S. bodinieri (Figure 10) in the posture, the rotundate or subrotundate leaf-lamina, the glabrous ovaries, and the pappose achenes, but can be distinguished by the smaller stature (10-12 vs. 10-35 cm), the smaller leaf-lamina (0.8-1.7 x 0.8-2 vs. 2-6 x 3-6 cm) shallowly regularly 5-7-lobed (vs. repand or
|
|
|
|
|
Figure 9. Geographical variation of leaf-lamina shape and hairiness in the Senecio bodinieri group. Triangles (▲) indicate populations with the leaf-lamina broadly ovate in outline and long-ciliate along the margin, and dots (•) indicate populations with the leaf-lamina rotundate or subrotundate in outline and short-ciliate along the margin.
|
|
|
|
|
|
|
|
Table 1. Comparison of Sinosenecio nanchuanicus and S. bodinieri (sensu Jeffrey and Chen).
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Lamina rotundate or subrotundate in outline, shallowly regularly 5-7-lobed with broadly deltoid or rounded lobules
|
Lamina rotundate or subrotundate in outline, repand or irregularly lobate with deltoid to broadly deltoid teeth or lobes
|
|
|
|
|
|
|
|
|
|
|
Lamina dark-green, pubescent above, purple beneath, glabrescent or sparsely pubescent on the midvein beneath, veins sunken above
|
Lamina green, rarely dark-green, sparsely
adpressed-pilose or glabrous above, pale green or purplish beneath, sparsely pubescent on the veins beneath, veins not sunken above
|
|
|
|
Anther endothecial cell wall thickenings
|
|
|
|
|
|
|
Constantly solitary and terminal
|
Several in terminal corymbs, rarely solitary
|
|
|
|
Ray florets per capitulum
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
On rock cliff under waterfall or along streams,
1,200-1,800 m a.s.l
|
Hillside, streamside or in woods, 650-2,700 m a.s.l
|
|
|
|
Geographical distribution
|
Restricted to Nanchuan County, Chongqing
|
Widely distributed in Chongqing, Guizhou, Hunan, Sichuan, and Yunnan
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Botanical Studies, Vol. 52, 2011
|
|
|
|
|
|
|
|
|
|
|
|
|
Figure 11. Sinosenecio palmatilobus (Kitam.) C. Jeffrey & Y. L. Chen (From Malinghe, Xingyi County, Guizhou, China and vouched by Qin-er Yang & Qiong Yuan 1078, IBSC).
|
|
|
|
in the community. Although the flowering period of S. bodinieri (April to June) partly coincided with that of S. nanchuanicus (June), we did not observe any possible interspecific hybrids at the site. In view of their remarkably different chromosome numbers, it is tempting to suggest that the difference in ploidy level may have effectively blocked their hybridization even when they grew together within a community.
The Jinfoshan Mountain is an important limestone karst area for biodiversity conservation, where Cathaya argyrophylla Chun & Kuang, a rare and endangered relict gymnosperm species endemic to China, has been recorded to occur. Notably, seven taxa of Sinosenecio have been discovered from this mountain: S. bodinieri (Vaniot) B. Nord., S. fanjingshanicus C. Jeffrey & Y L. Chen, S. glo-bigerus (Chang) B. Nord. var. adenophyllus C. Jeffrey & Y. L. Chen, S. oldhamianus (Maxim.) B. Nord., S. septilobus (Chang) B. Nord., S. subcoriaceus C. Jeffrey & Y. L. Chen and the new species S. nanchuanicus, with four of them, S. globigerus var. adenophyllus, S. septilobus, S. subco-riaceus and S. nanchuanicus being locally endemic to the area. This remarkable local endemism and species richness in limestone karst areas in southern China have also been noted in several other plant groups (Hou et al., 2009; Yuan and Yang, 2009), indicating that further botanical exploration may unveil many more novelties in such groups.
Acknowledgments. We are very grateful to Dr. B. Nor-denstam for his invaluable comments on the manuscript. Our thanks are due to the director of E for the loan of specimens, to Ms. Yun-xiao Liu for making the drawing, and to Mr. Ming-bo Ren, Mr. Xiang Liu and Mr. Tao Deng for help in the field. This work was supported by the Knowledge Innovation Project of the Chinese Academy of Sciences (KZCX2-YW-415, KSCX2-YW-Z-0918) and the National Natural Science Foundation of China (Grant no.
30970183).
|
|
|
|
Figure 10. Sinosenecio bodinieri (Vaniot) B. Nord. (sensu Jeffrey and Chen herein). A, Posture; B, Capitula (above) and florets (below); C, Leaf (above: adaxial side; below: abaxial side); D, Habitat (All from Jinfoshan, Nanchuan, Chongqing, China and vouched by Ying Liu & Tao Deng 2008042, IBSC, PE).
|
|
|
|
irregularly lobate with deltoid to broadly deltoid teeth or lobes) and dark-green above (vs. usually green), the veins sunken above (vs. not sunken), the capitula constantly solitary and terminal (vs. usually several in corymbs, very rarely solitary), and the rays 14-15 (vs. 13). From S. palmatilobus (Figures 7C, D, and 11), the new species differs also by these characters and, additionally, by its leaf-lamina rotundate or subrotundate in outline (vs. broadly ovate) and short-ciliate along the margin (vs. long-ciliate). The recognition of this new species is strongly supported by chromosomal data, with S. nanchuanicus being a hexaploid with 2n = 144 while its putative close relative, S. bodinieri, being a diploid with 2n = 48. The chromosome number of S. palmatilobus, counted from a population from Xingyi, Guizhou, has also 2n = 48 (Ying Liu & Qin-er Yang, unpublished data).
In the Jinfoshan Mountain area, the type locality of our new species Sinosenecio nanchuanicus, S. bodinieri is very common, whereas the new species is rather rare. In the field, we noticed that the latter seldom flowered, with most of its individuals being maintained at vegetative stage. Interestingly, at one site, we found both species grew together on a rock cliff under a small waterfall, with S. bodinieri dominant while S. nanchuanicus quite rare
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
LIU et al. ― Sinosenecio nanchuanicus, a new species from China
|
|
|
|
|
|
|
|
|
|
thesis, Institute of Botany, Chinese Academy of Sciences, Beijing.
Liu, Y and Q.E. Yang. 2010. Sinosenecioyilingii (Asteraceae), a new species from Sichuan, China. Bot. Stud. 51: 269-275.
Liu, Y., G.X. Chen, and Q.E. Yang. 2009. Sinosenecio baojin-gensis (Asteraceae), a new species from Hunan, China. Bot. Stud. 50: 107-113.
Liu, Y., D.G. Zhang, and Q.E. Yang. 2010. Sinosenecio hup-ingshanensis (Asteraceae), a new species from Hunan and Hubei, China. Bot. Stud. 51: 387-394.
Nordenstam, B. 1978. Taxonomic studies on the tribe Sene-cioneae (Compositae). Opera Bot. 44: 1-84.
Zhang, D.G., Y. Liu, and Q.E. Yang. 2008. Sinosenecio jishouen-sis (Compositae), a new species from north-west Hunan, China. Bot. Stud. 49: 287-294.
Yuan, Q. and Q.E. Yang. 2009. Anemone xingyiensis (Ranuncu-laceae), a new species from Guizhou, China. Bot. Stud. 50:493-498.
Vaniot, M. 1902. Composees. Bull. Acad. Geogr. Bot. 11: 345351.
|
|
|
|
Chen, Y.L. 1999. Sinosenecio B. Nord. In Anonymous (ed.), Flora Reipublicae Popularis Sinicae 77(1). Science Press, Beijing, pp. 101-141.
Jeffrey, C. and Y.L. Chen. 1984. Taxonomic studies on the tribe Senecioneae (Compositae) of Eastern Asia. Kew Bull. 39:205-446.
Handel-Mazzetti H. 1937. Neue und bemerkenswerte chinesi-sche Compositen, besonders aus dem Berliner Herbar. No-tizbl. Bot. Gart. Berlin. 13: 607-661.
Hou, M.F., Y. Liu, Y. Kono, and C.-I Peng. 2009. Aspidistra dax-inensis (Ruscaceae), a new species from limestone areas in Guangxi, China. Bot. Stud. 50: 371-378.
Kitamura, S. 1982. Compositae Asiaticae. Acta Phytotax. Geo-bot. 33: 190-200.
Lauener, L.A. 1976. Catalogue of the names published by Hector Leveille: IX (Compositae). Notes R.B.G. Edinb. 34: 327 -402.
Liu, J.Q. 1999. Systematics of the Tribe Senecioneae subtribe Tussilagininae (Asteraceae) of the Eastern Asia. Ph.D.
|
|
|
|
|
|
|
|
中國重慶產蒲兒根屬一體型小而染色體數目高的新種:
南川蒲兒根
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1中國科學院植物資源保護與可持續利用重點實驗室 (華南植物園)
2重慶巿藥物種植研究所
3中國科學院植物研究所系統與進化植物學國家重點實驗室
|
|
|
|
|
|
|
|
本文描述了中國重慶產蒲兒根屬一新種:南川蒲兒根(Sinosenecio nanchuanicus Z. Y. Liu, Y. Liu &
Q. E. Yang)。其體細胞染色體數目為2n = 144 。本新種在體態,葉片圓形或近圓形,瘦果具冠毛方面
與染色體數目為2n = 48的滇黔蒲兒根(S. bodinieri (Vaniot) B. Nord.)最為相似,但除染色體數目不同
外,還以植株較小,葉較小,葉片上表面深藍綠色,葉脈下陷,葉5-7規則淺裂,頭狀花序單生,舌狀
花14-15而與後者相區別。本新種也以同樣的特徵以及葉片圓形或近圓形、邊緣具較長緣毛而與掌裂蒲
兒根(S. palmatilobus (Kitam.) C. Jeffrey & Y. L. Chen)相區別。本文提供了南川蒲兒根以及滇黔蒲兒根
和掌裂蒲兒根的彩色圖版和一些標本照片、南川蒲兒根的線繪圖、花部微觀性狀的光鏡照片以及地理分
佈圖。
|
|
|
|
|
|
|
|
關鍵詞:菊科;染色體數目;石灰岩植物區系;地方特有現象;多倍體;千里光族;南川蒲兒根。
|
|
|
|
|
|
|
|
|
|
|
|