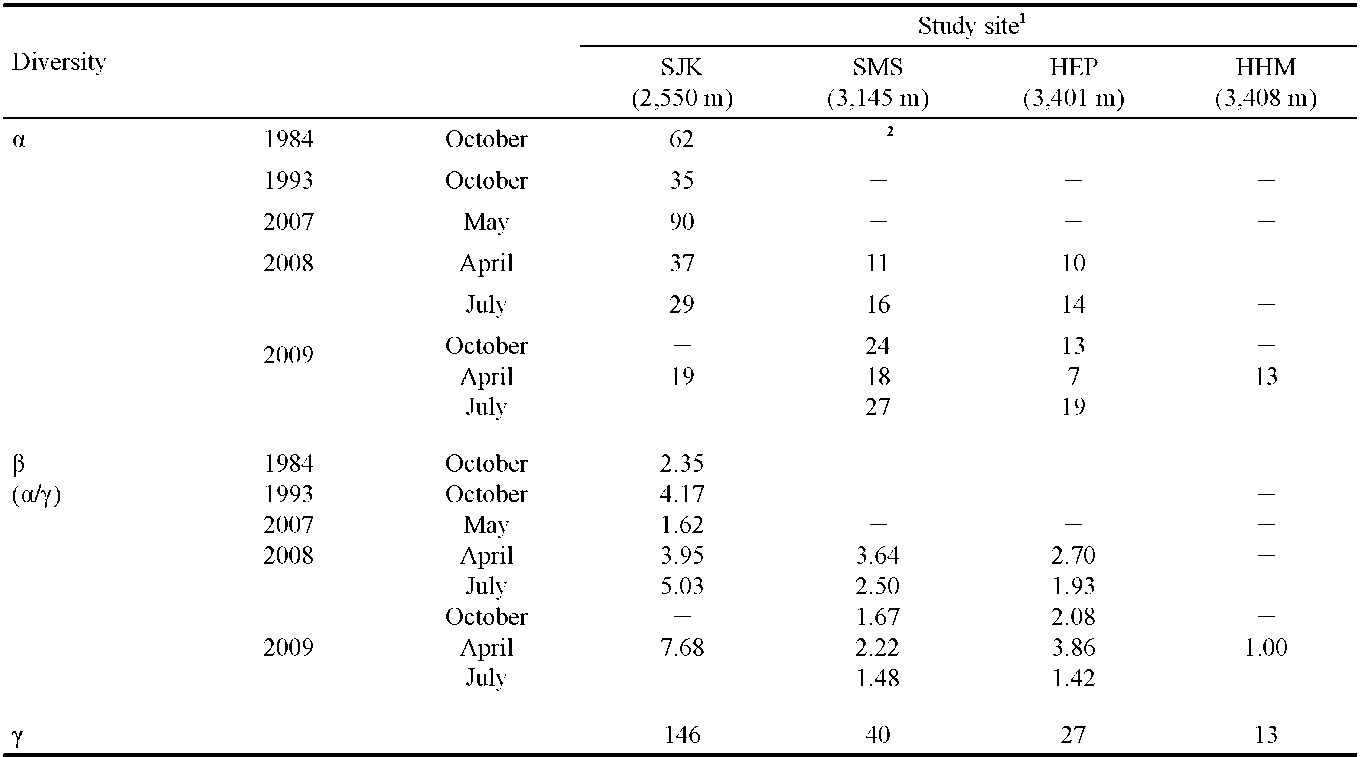CHOU et al. ―Diversity of alpine vegetation affected by climate change
505
ntu.tw/webtaiprog/web.query-aspx), and Taiwan Forestry Research Institute (http://taif.tfri.gov.tw:8080/spec-query.aspx). We surveyed plant species distributed in the He-huanshan area, arbitrarily divided into 4 elevation zones (2,000-2,500 m, 2,500-3,000 m, 3,000-3,500 m, and above 3,500 m) and listed the plant species, genera, and families found within each zone. The plant species nomenclature follows Flora of Taiwan (2nd edition, Huang et al., 2002); in addition, a list of rare and endangered species was ob-tained from the database of the Council of Agriculture, Taiwan (Chou et al., 2009, unpublished report).
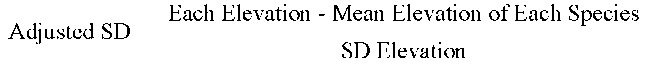
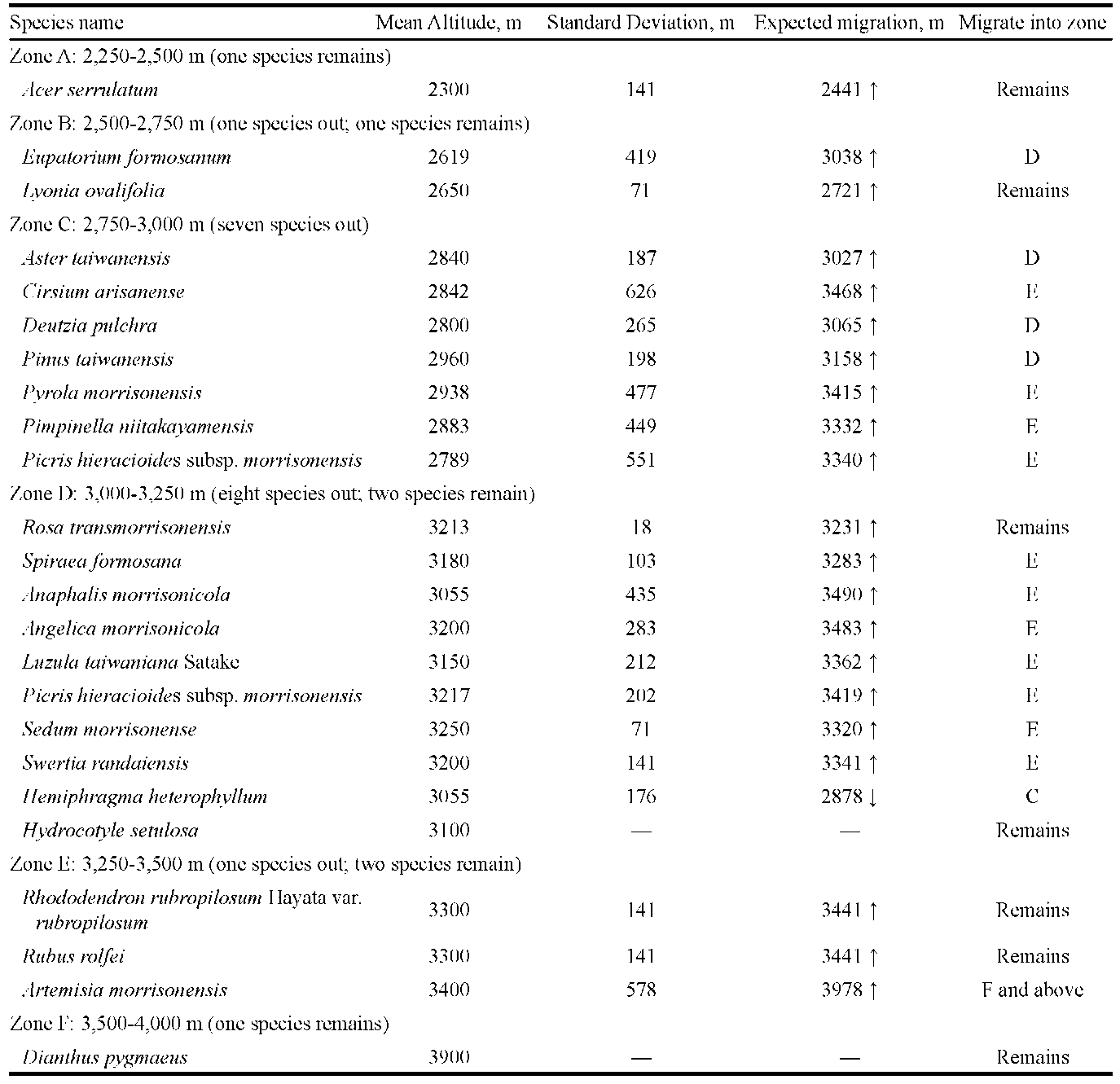
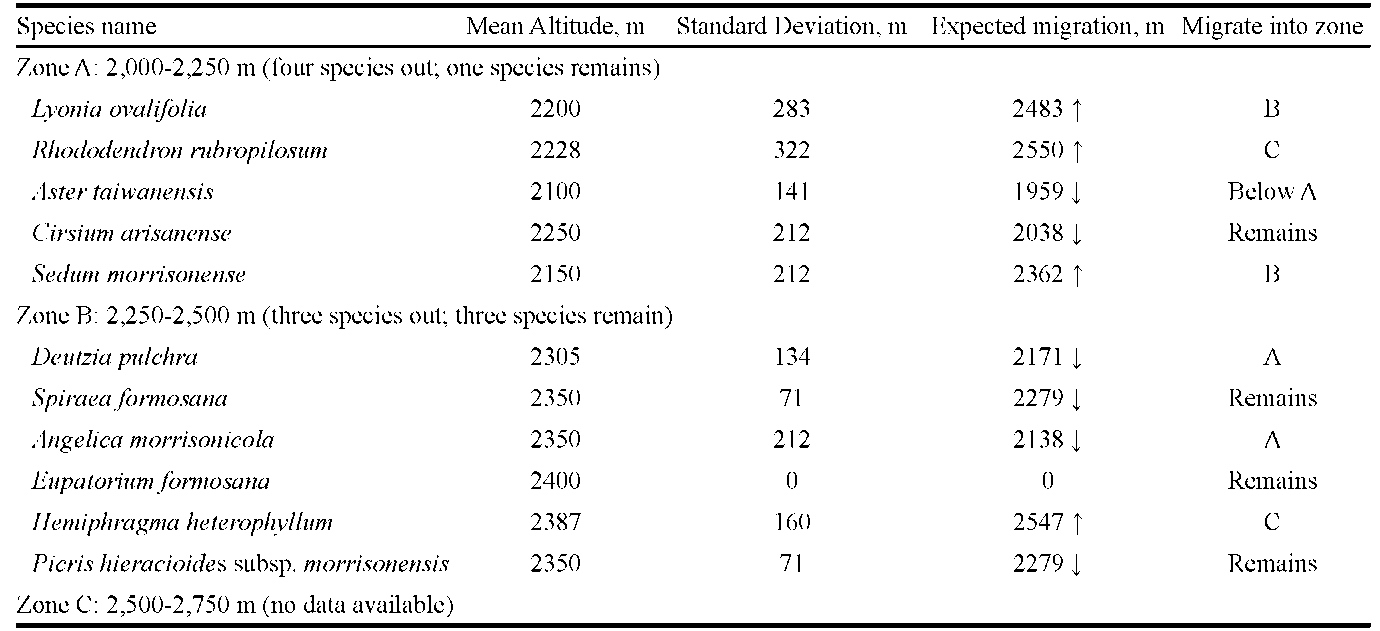
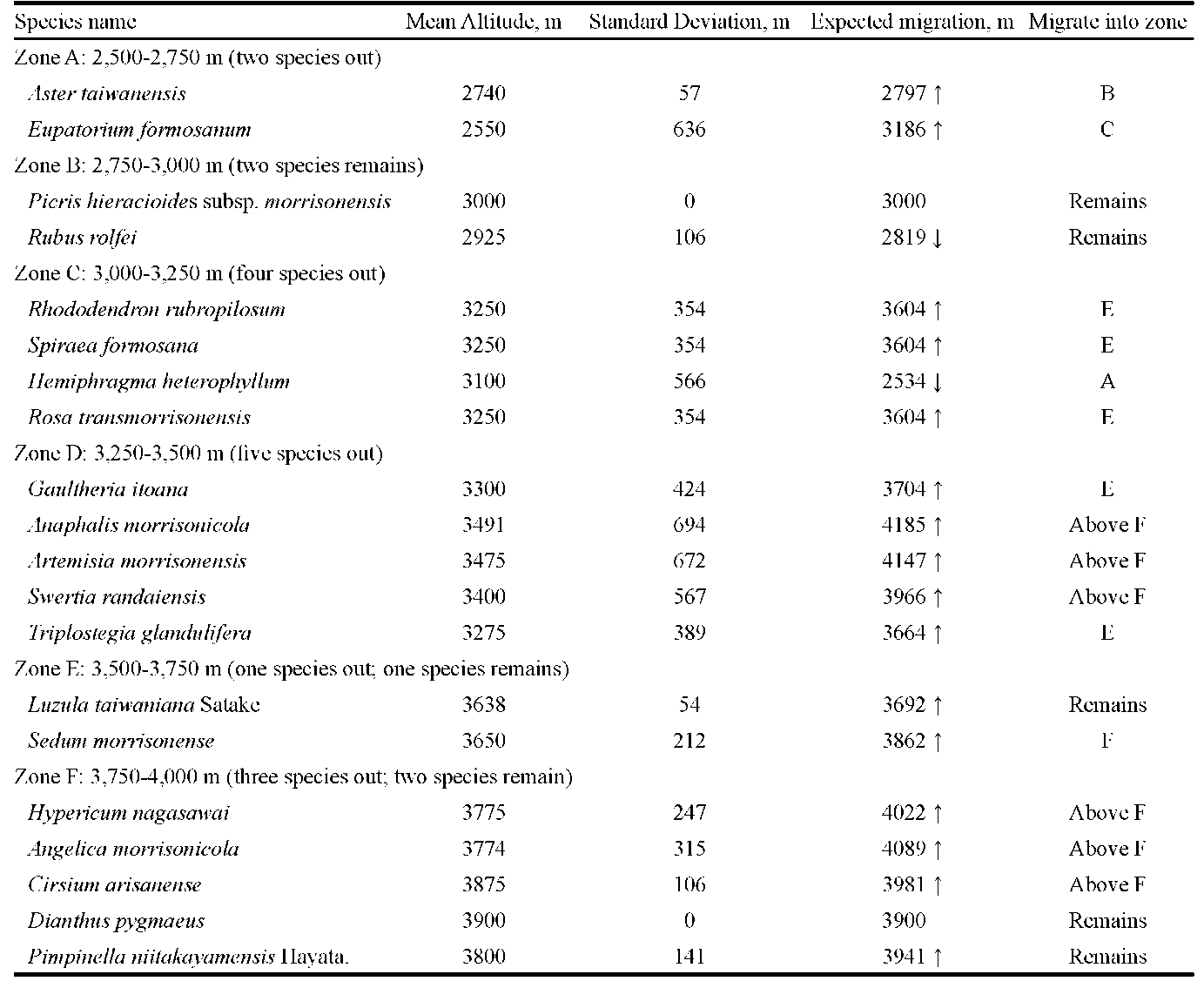
The expected elevation of migrated species was represented by M (mean elevation in meter) plus SD (standard deviation). For example, Acer serrulatum was documented at the first time in 1984 at 2,200 m asl and was later documented in 1999 at 2,400 m asl. From the two data, we obtained a mean elevation of 2,300 m and a standard deviation (SD) of 141.42 m. From the mean and SD, it was deduced that the plant might move upwards to an expected elevation of 2,441 m. Following this computing process, we were able to assess the migration of each plant to a certain altitude.
Diversity measurement
We determined the a, p, and y diversity of vegetation and used Simpson's diversity index to make a comparison between sampling sites and time. The equation of Simpson's diversity index was described by Simpson (1949).
Estimation of available land area in the Hehuan-shan area
At the study sites, plants were sampled based on the botanical survey of quadrates (10 x 10 m each) and on and transect sampling (20 m long) at an interval of 100 m along Highway 14A from an elevation of 2,250 to 2,750 m.
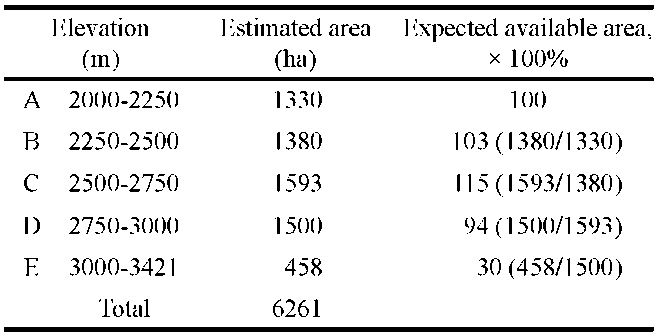
Using ArcView software and a digital geographic model (Digital Terrain Model, DTM), and through communication with Prof. Chen at the National Pingtung University of Science and Technology, Taiwan, the land area of Hehuanshan above 2,000 m was estimated by dividing into five elevational zones (2,000-2,250 m, 2,250-2,500 m, 2,500-2,750 m, 2,750-3,000 m, and above 3,000 m). The estimated available land area is expressed as a percentage (Chou et al., 2009, unpublished report).
Examination of plant specimens in herbaria in Japan and Taiwan
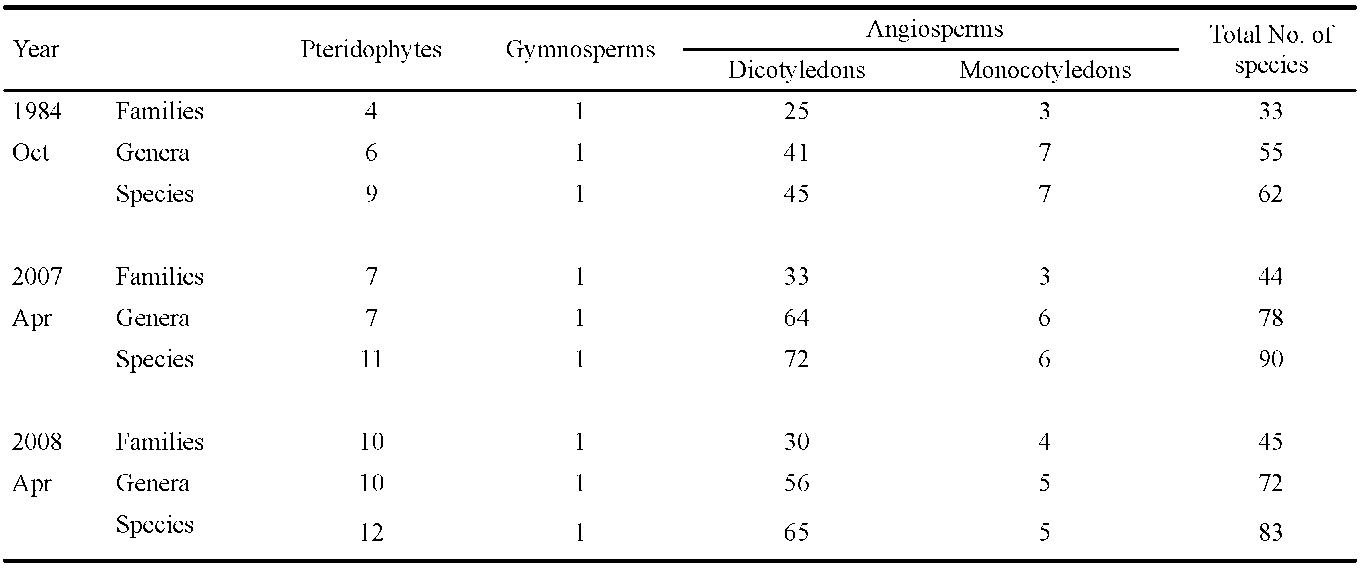
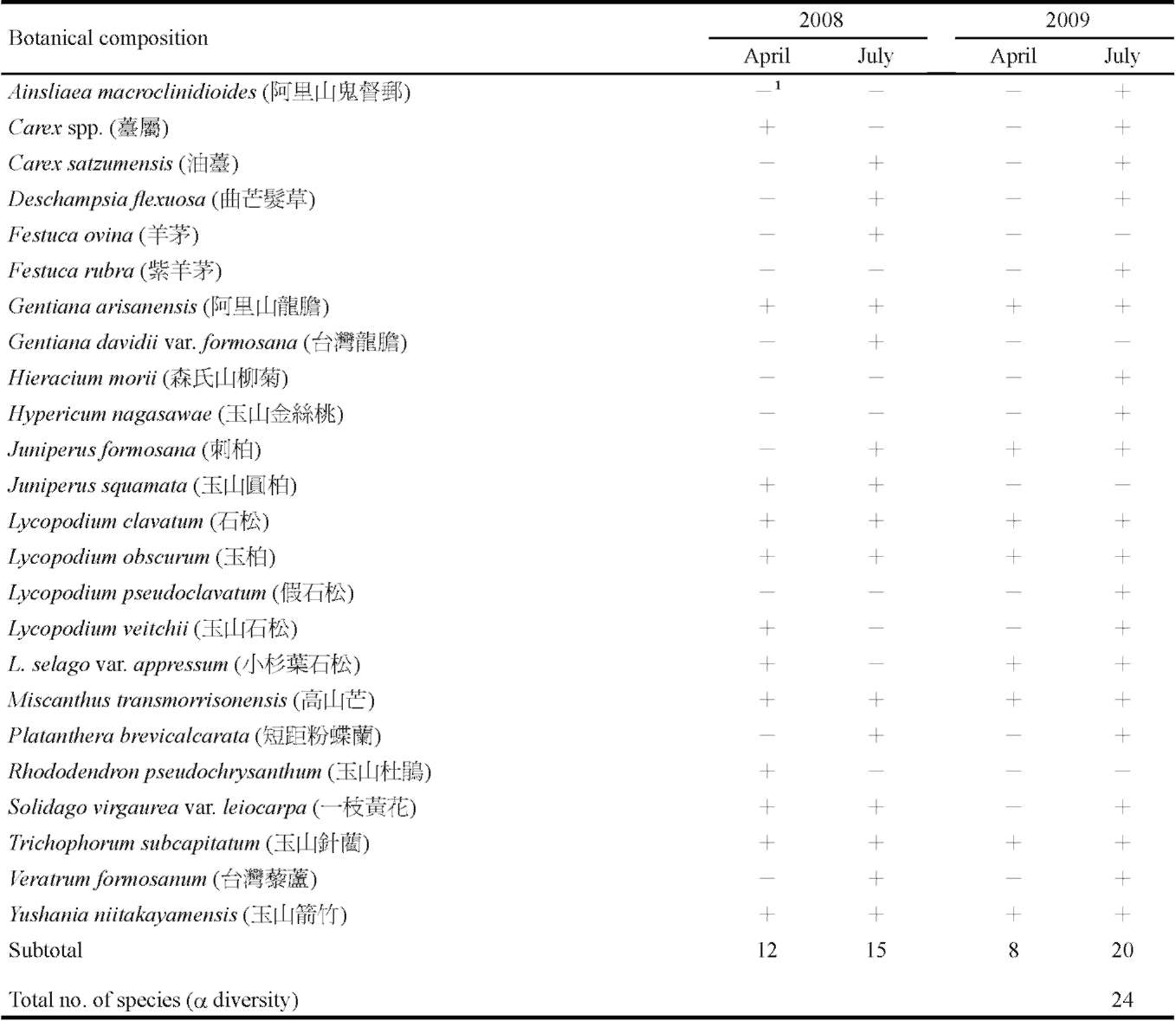
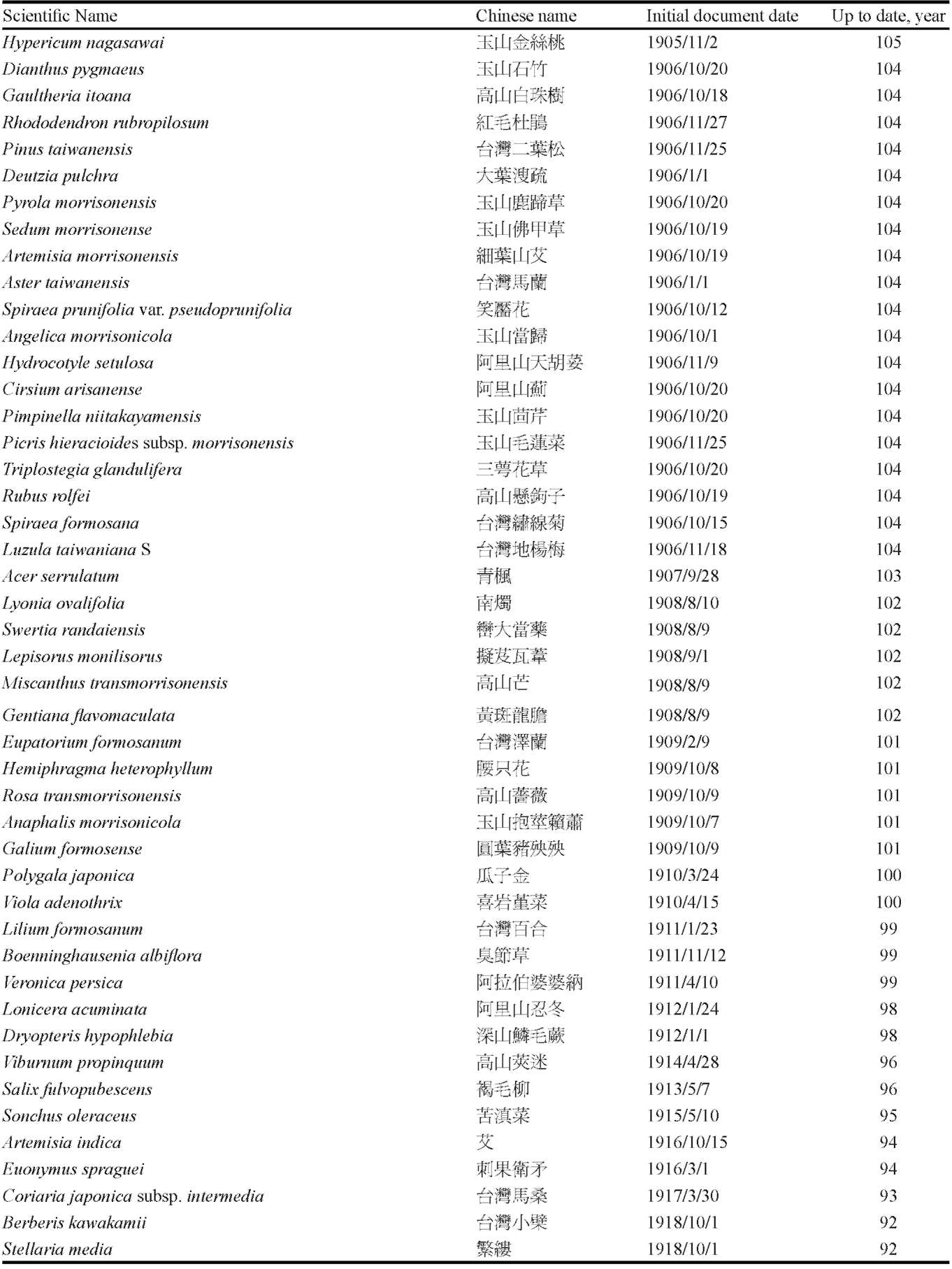
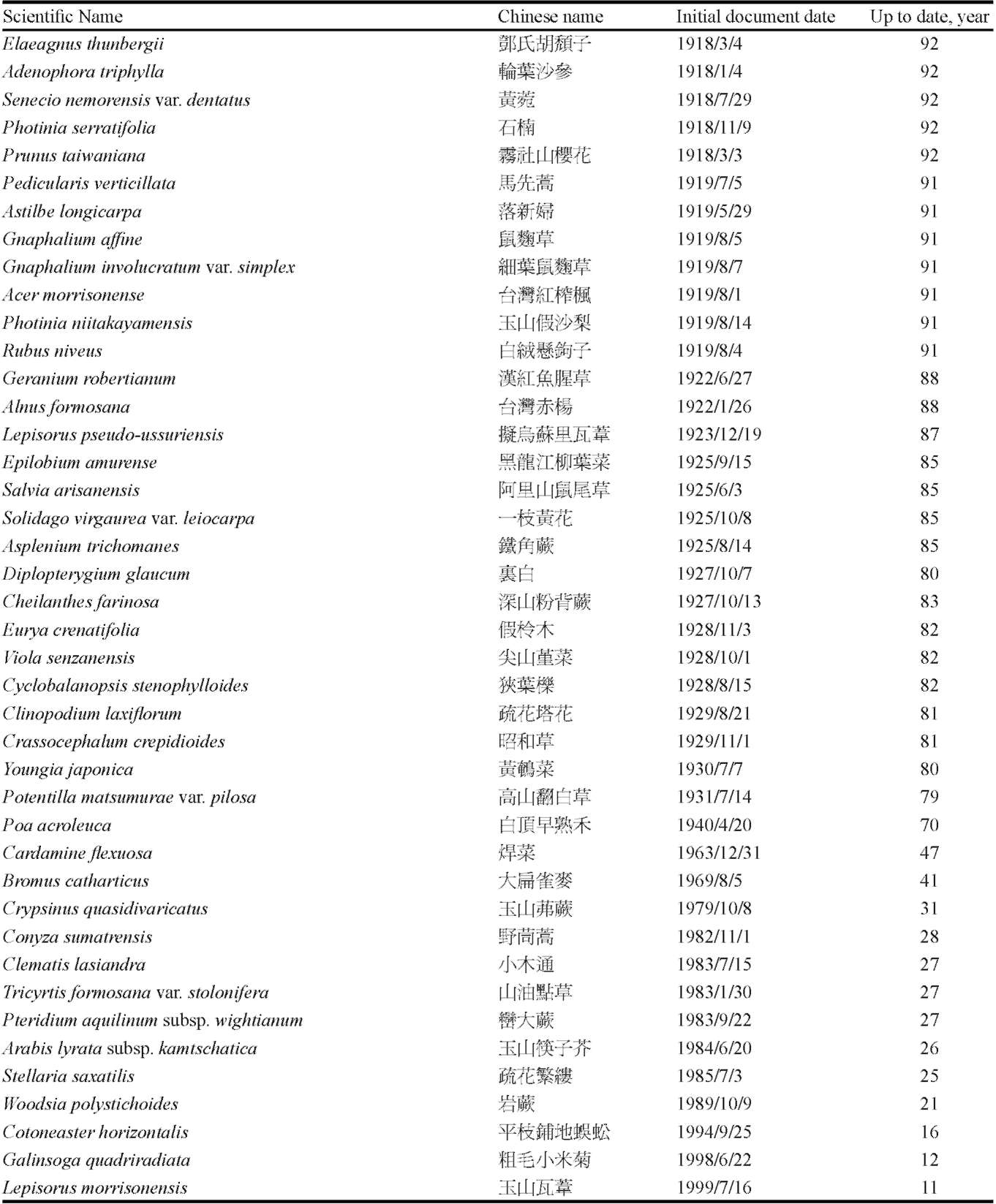
From the databases mentioned above, we downloaded relevant information for each species found at our selected study sites. We consulted the herbarium of Tokyo University to add any information from early documents and recent findings that had not been uploaded to the databases. For the 367 plant species that were recorded in the Hehuanshan area, we carefully checked all vouchers deposited in the herbaria and the contents of several databases in Taiwan. We obtained a list of species found at the Shinjenkan site in different years and at different elevations (Chou et al. 2009, unpublished report, or see Supplementary Table 1).
RESULT AND DISCUSSION
Botanical diversity in the alpine area of Taiwan
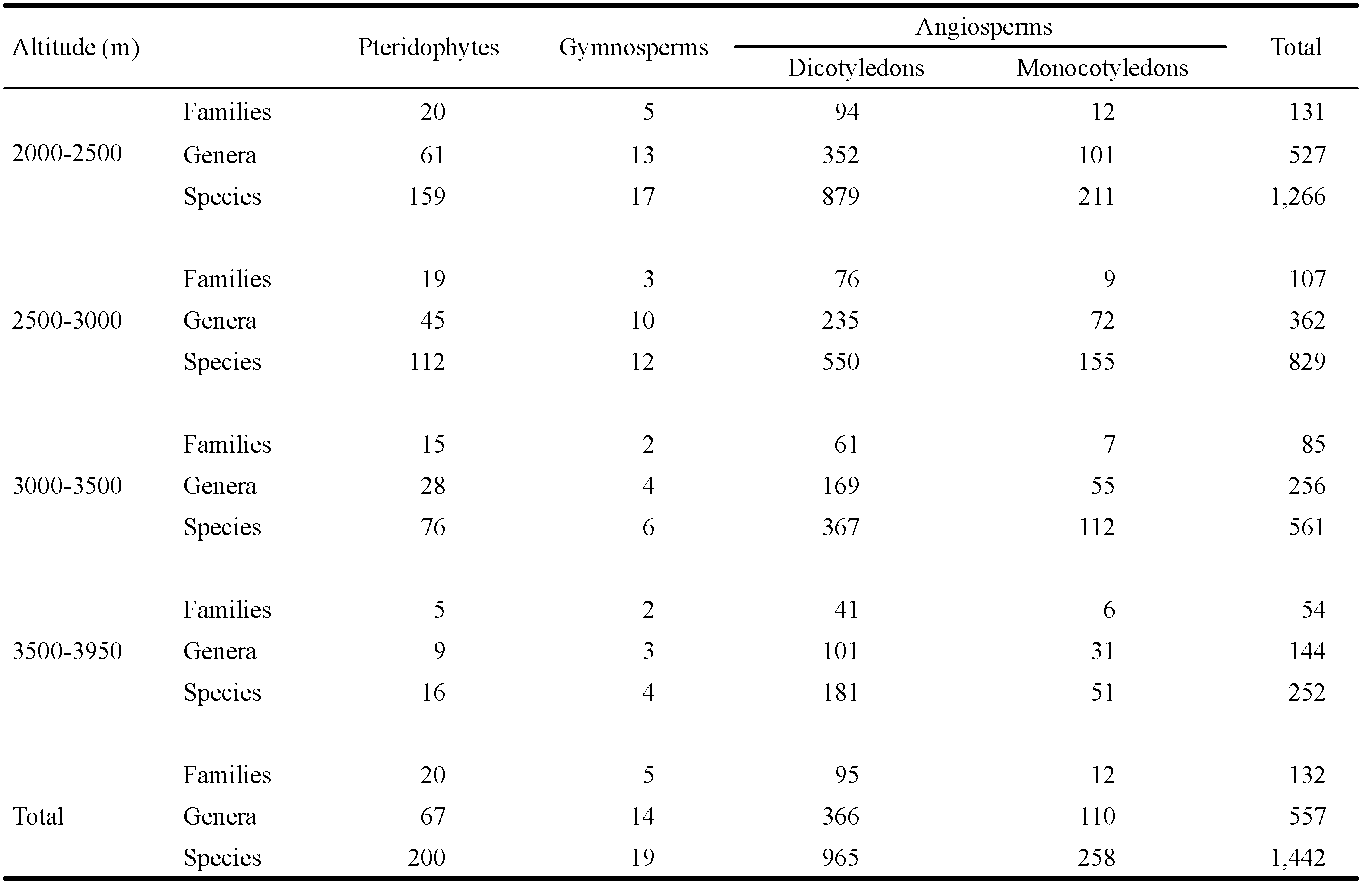
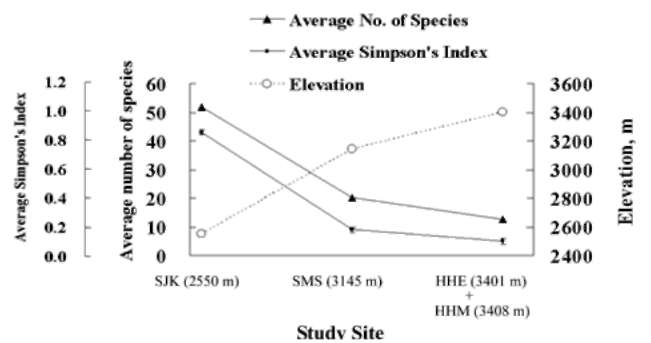
We attempted to study the botanical diversity of alpine plants based on a century of botanical inventories documented in the databases of various Taiwanese institutions. Appropriately 25,000 specimens have been collected in our study area above an elevation of 2,000 m, including 132 families, 557 genera, and 1442 species (Table 1). In the elevation zone of 2,000-2,500 m, we recorded 1266 species, 527 genera, and 131 families; in the 2,500-3,000 zone we recorded 829 species; in the 3,000-3,500 m zone we recorded 561 species; and above 3,500 m we recorded 252 species (Huang et al., 2002). It is clear that the number of plant species decreases with increasing elevation.
Checking climatic data from the last century in Taiwan
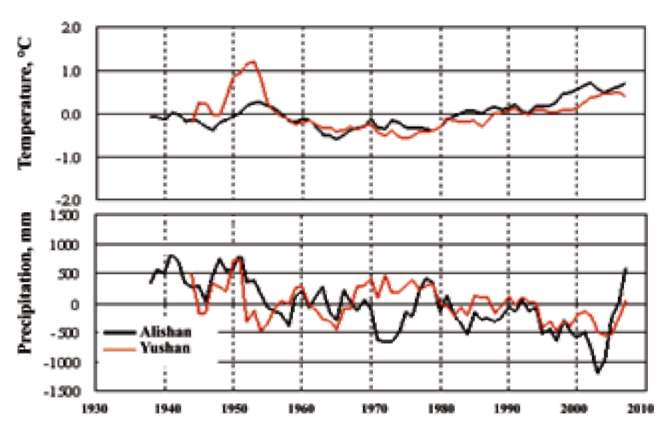
Since 1895, weather stations have been established in most areas in Taiwan. Weather data from the last century were obtained from the Central Weather Bureau of Taiwan; however, the data from the Hehuanshan are incomplete due to the lack of a weather station there at the beginning of the 20th century. Instead, we obtained data from nearby stations at Alishan and Yushan, which are relatively close to the Hehuanshan site. The average annual temperature and precipitation since 1930 are presented in Figure 1.
At the same time, the available land decreases with altitude. For example, in the Hehuanshan area, the available land in elevation zone C (2,500-2,750 m) corresponds to 94% of the land area in elevation zone D (2,750-3,000 m), and the area in zone E (3,000-3,421 m) is equal to 30% of the area in elevation zone D (2,750-3,000 m) (Table 2). Logistically, under global warming in the region, plant species would migrate upwards to the higher elevations in zone E, where the available land for plant growth is only 30% of the area previously occupied (Table 2); in other words, about 70% of plant species might not survive due to limited space. However, most endemic species may not be seriously affected by global warming. In the Hehuan-shan area, there are 480 endemic species, about one-third of the total number of species found in the mountainous
Statistical analysis
All species are listed in an Office Excel 2007 table according to altitude, and the elevations of each species' distribution vs. year of each species' recording were analyzed statistically using standard deviation (SD) as described by Gomez and Gomez (1976). An adjusted SD was obtained according to the formula which was used by Jump et al. (2006):