68
Botanical Studies, Vol. 53, 2012
structure of maize (Ciamporova, 2000). At the cellular level, the possible sites of Al effects occurring are in the cell wall, on the plasma membrane or in the cytoplasm. Possible mechanisms of Al toxicity involve Al interactions with the cell wall constituents and the plasma membrane (Delhaize and Ryan, 1995; Horst, 1995).
iments. The experimental results were submitted to ANOVA of simple classification. The statistical package STAT-GRAPHICS (version 4.1 for Windows) was used to calculate both the SE and to compare the means (Tukey's test).
RESULTS
During the past few decades, physiologists have contributed
to our understanding of the mechanisms of Al
toxicity and tolerance in some of the cereals (Matsumoto,
2000; Kochian et al., 2005; Zheng and Yang, 2005; Rubia
et al., 2011). Although rice is generally considered the
most Al-tolerant species among small grain cereal crops,
the mechanisms responsible for the high Al tolerance of
rice are not yet understood (Macedo and Jan, 2008).
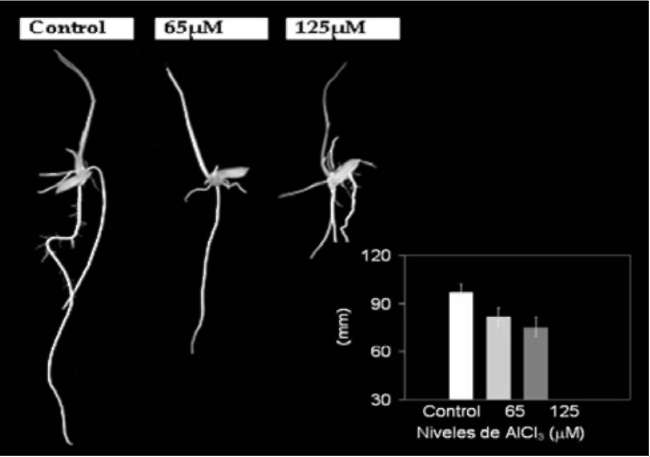
Root elongation was significantly inhibited in rice seedlings
exposed to Al (Figure 1). However, growth inhibition
was more severe in seedlings exposed to 125 μM than in
seedlings exposed to 65 μM AlCl3. In the most stressed
roots, elongation was reduced 24% compared to the control
plants, while those exposed to 65 μM AlCl3 exhibited
a reduction of 15.5% (Figure 1).
The aim of this paper is to study the responses to Al to-xicity in a Cuban rice cultivar (INCA LP-7). The effects of Al on root morphology and on some cellular level changes are also investigated.
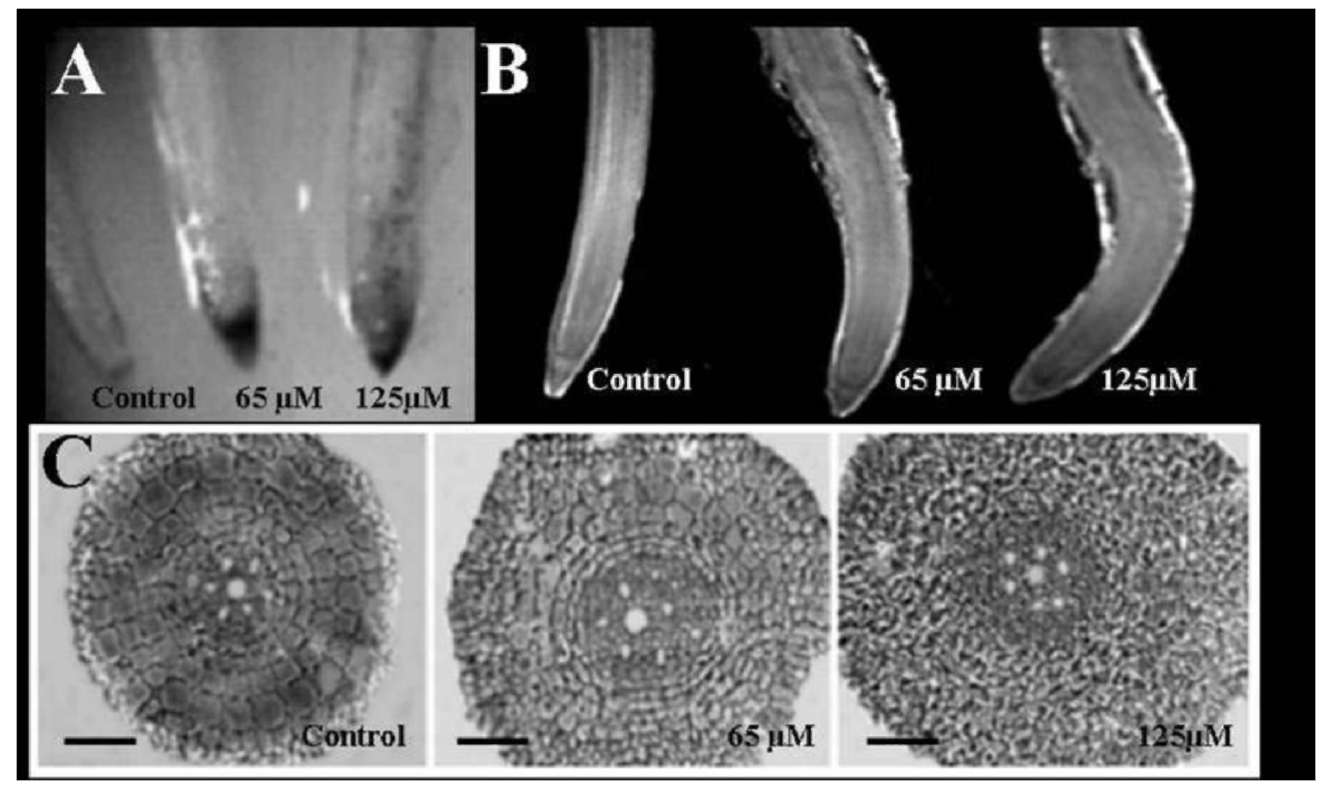
Seminal roots stained with hematoxylin are shown in
Figure 2A. In seedlings exposed to Al, the root tips exhibited
a red-brown staining, which was more intensive in
roots exposed to 125 than to 65 μM Al. Seedlings exposed
to 125 μM also exhibited stained spotting. No staining was
observed in control plants (Figure 2A).
MATERIALS AND METHODS
Plant material and growth conditions
Roots of stressed seedlings showed symptoms of Al injury at both Al concentrations. Root-apices appeared swollen and irregularly curved in Al-treated seedlings (Figure 2B). A localized thickened zone was observed between 20 and 40 mm from the rice root apex, corresponding to an increased diameter of cross-sections (Figure 2C)
Seeds of Oryza sativa L., cv INCA-LP7 were sterilized with 5% sodium hypochlorite and germinated on filter paper for seven days. Two Al concentrations were employed to impose the stress; 0, 65 and 125 (iM AlClg (pH= 4). The seedlings were under 16 h of light/8 h of darkness at 25°C.
To evaluate root elongation, the primary roots of seedlings (n > 25) were photographed and their length measured with a ruler before microscopic sampling.
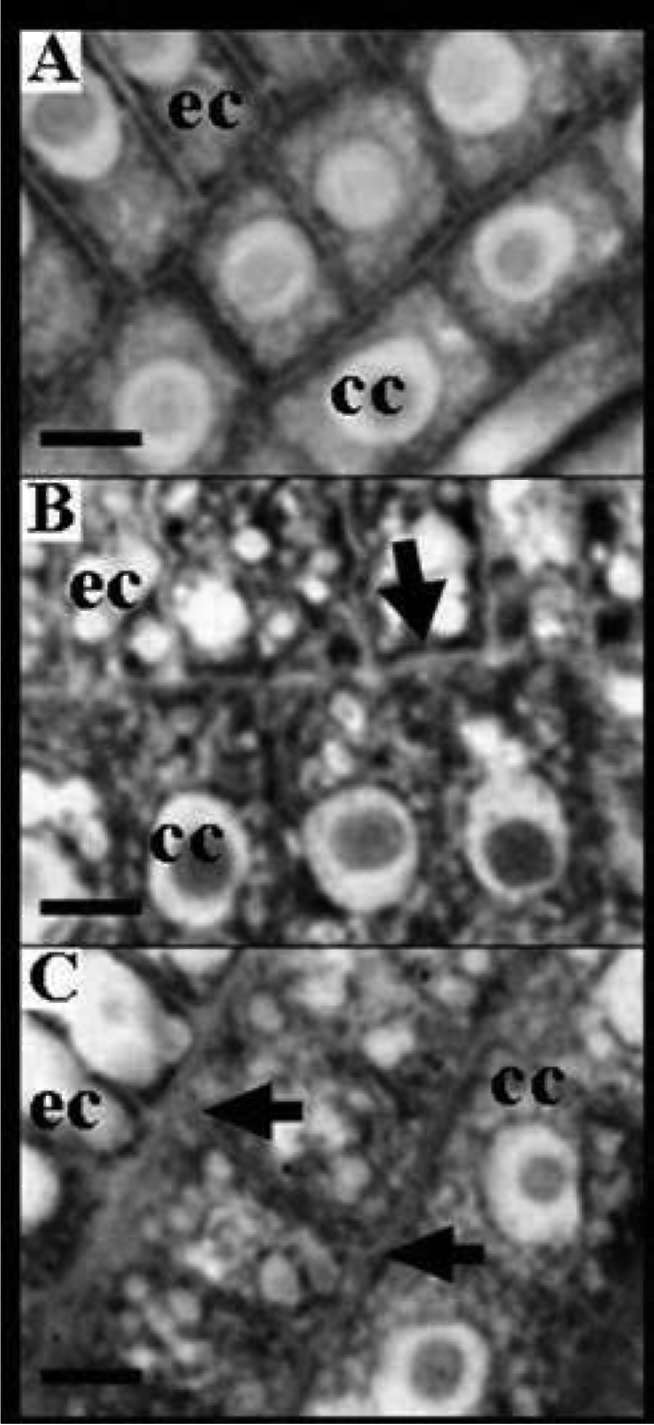
Cross-sections of roots exposed to both Al concentrations
were observed as more disordered (Figure 2C) than
those of the control roots. Some alterations in the size of
epidermal and cortical cells provoked by Al were responsible
for this change. The epidermal cells were of typical
size in longitudinal sections of the control roots (Figure
3), but lost their tissue features and appeared shorter and
wider than the cells in control roots, when exposed to 65
and 125 μM AlCl3. These changes were also observed in
cortical cells (Figure 3).
Hematoxylin staining
Staining protocol was based on Polle et al. (1978). The roots of seedlings cultivated for seven days, in the presence or absence of Al, were gently shaken in 200 ml distilled water for 15 min. The water was then replaced by 200 ml of aqueous hematoxylin solution [0.2% hematoxylin (Merck) and 0.02% potassium iodide, w/v] and left at the same slow agitation for 20 min. The solution was then replaced with 200 ml of water. The root apices were excised and photographed under stereoscopic microscope.
Light microscopy
Apical segments obtained from primary roots (3 mm) were fixed with 4% paraformaldehyde. After three washes in distilled water, the roots were dehydrated by Progressive Lowering Methods (PLT) and embedded in Lowicryl resin (Risueno, 2000).
Semi-thin longitudinal and cross-sections were made on a LKB Ultramicrotome and stained with toluidine blue. The sections were viewed with a Zeiss optic microscope and photographed with a digital camera coupled to the microscope.
Figure 1. Effects of 65 and 125 (iM AICI3 on root elongation of rice cv INCA LP-7. Values are means of five independent replicates. Means followed by different letters differ significantly at P < 0.05 (Tukey's Test).
Statistical Analysis
Randomized complete design was employed for the exper-