106
Botanical Studies, Vol. 53, 2012
|
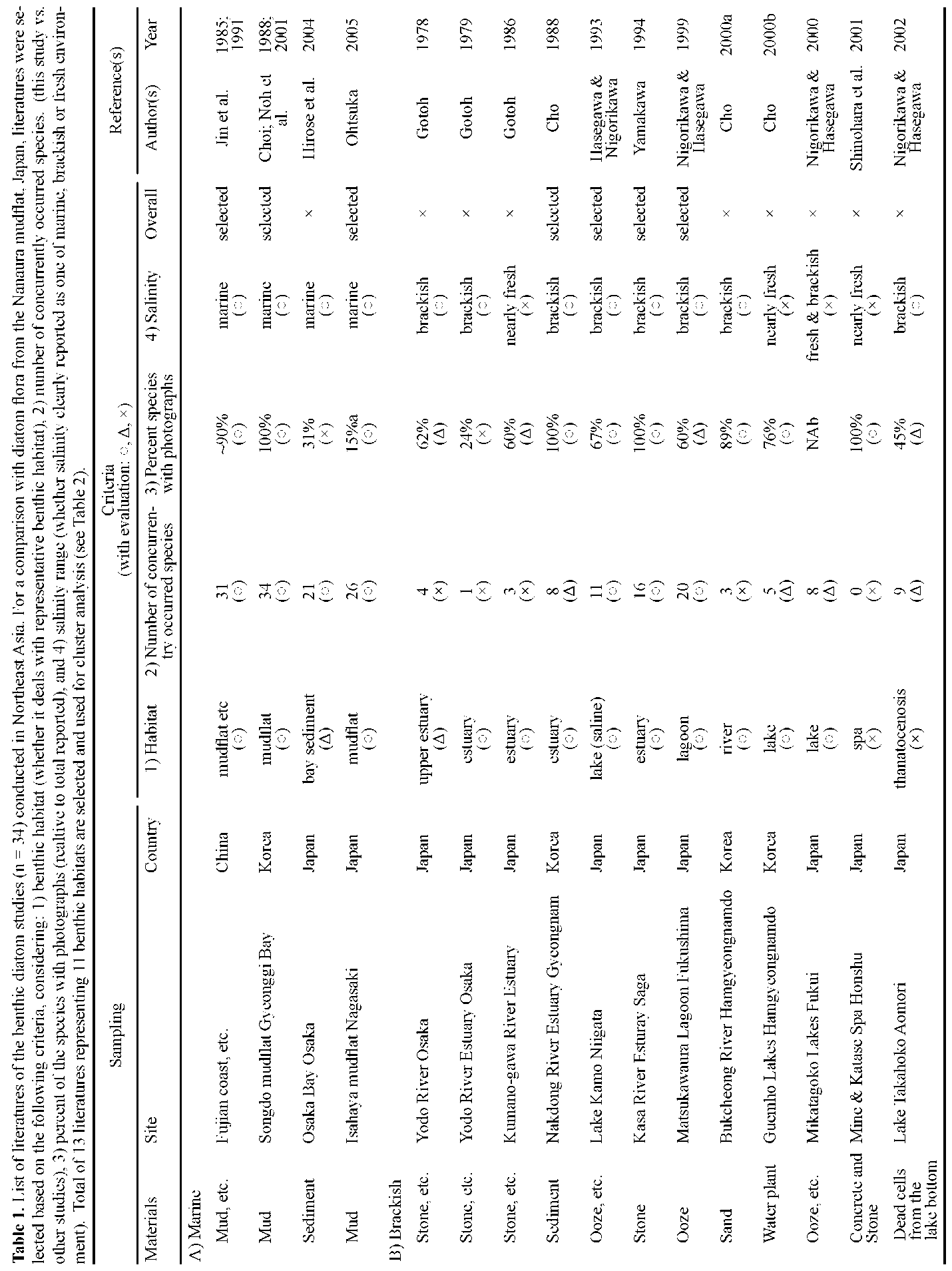
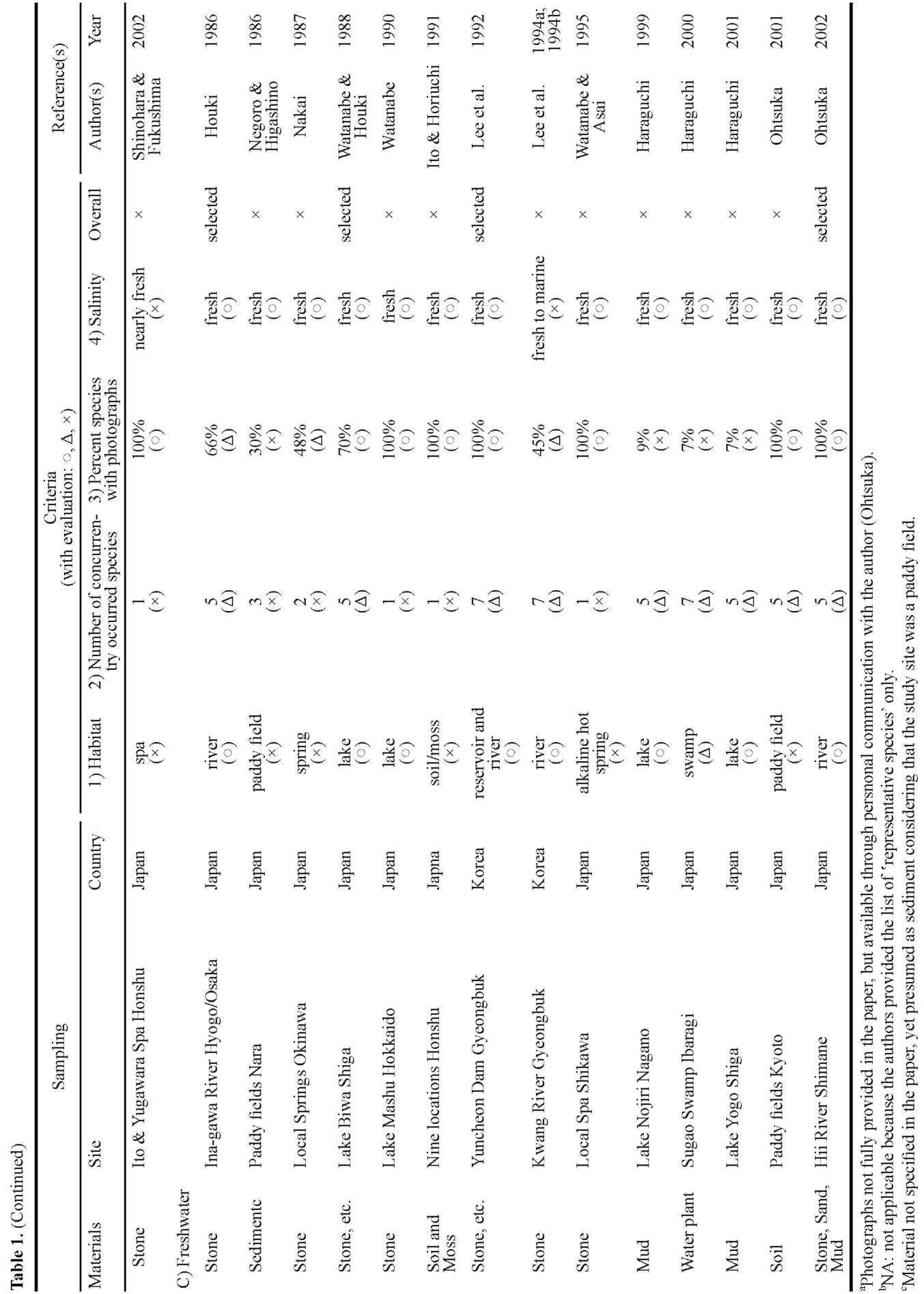
ports are available on marine benthic diatoms in the three countries listed above, all available literature (n=34) from Northeast Asia encompassing tidal flat or coastal (n=6), as well as other benthic environments, such as estuary (n=5), lake (n=7), river (n=6), and others (n=10), are included for comparison.
|
||
|
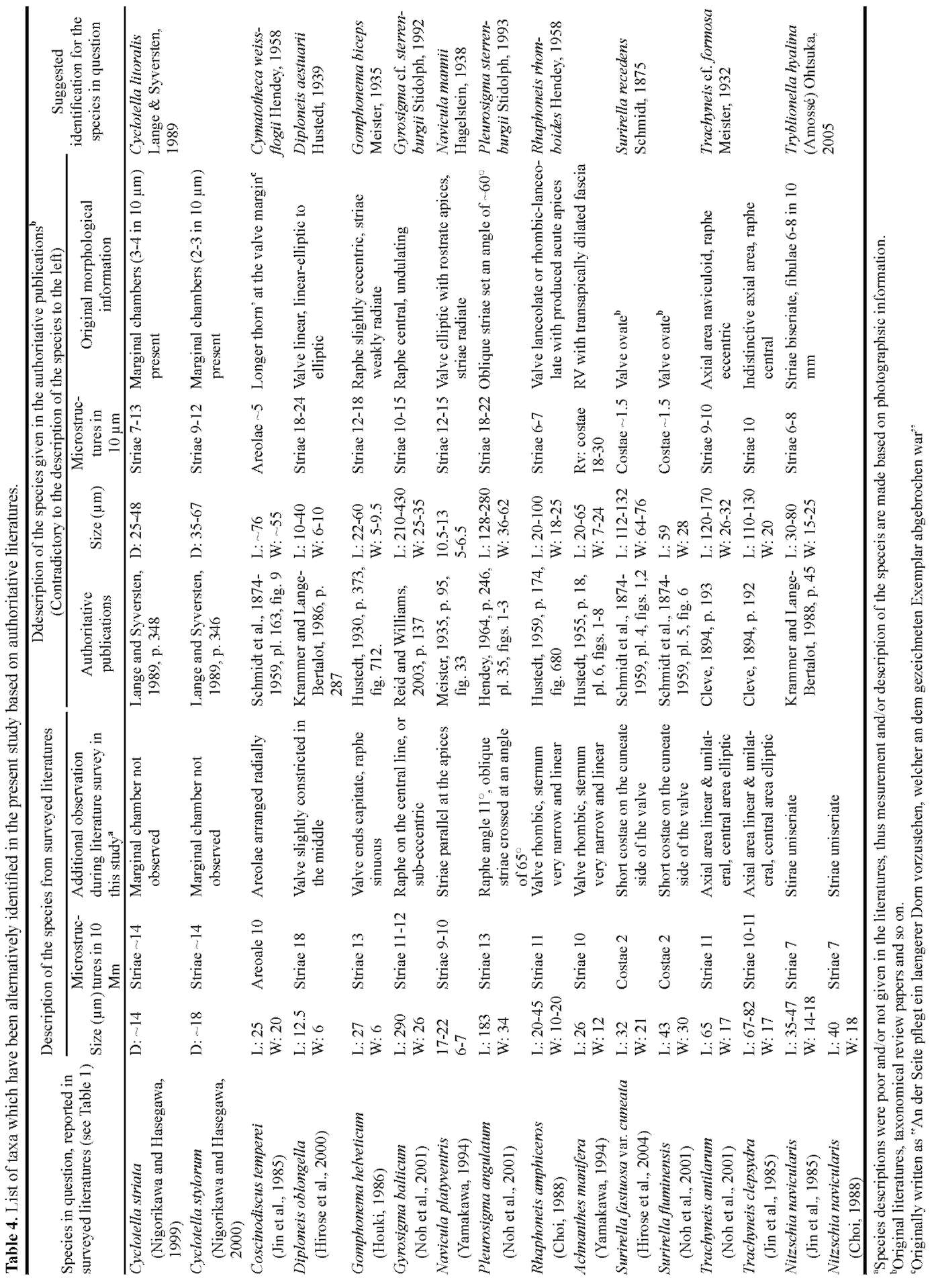
The nomenclature and flora established in Europe and North America have long been adopted and applied to flo-ristic and taxonomic studies on benthic diatoms in Asian countries. Using this approach, the taxa identified in the region were considered cosmopolitan. While few endemic forms were reported, their high occurrence in such unexplored habitats was expected. Thus, the correct identification of benthic diatoms in our materials and of other samples from Northeast Asia, is vital for floristic and taxo-nomic studies, especially for comparative studies. Comparisons based solely on checklists are misleading, and photographic documentation and morphological information should be addressed as far as possible. In the present study, those species likely misidentified and/or in question among the reviewed literature were carefully reevaluated. Alternative identifications were suggested accordingly and used for regional comparisons before larger comparisons were made.
|
||
|
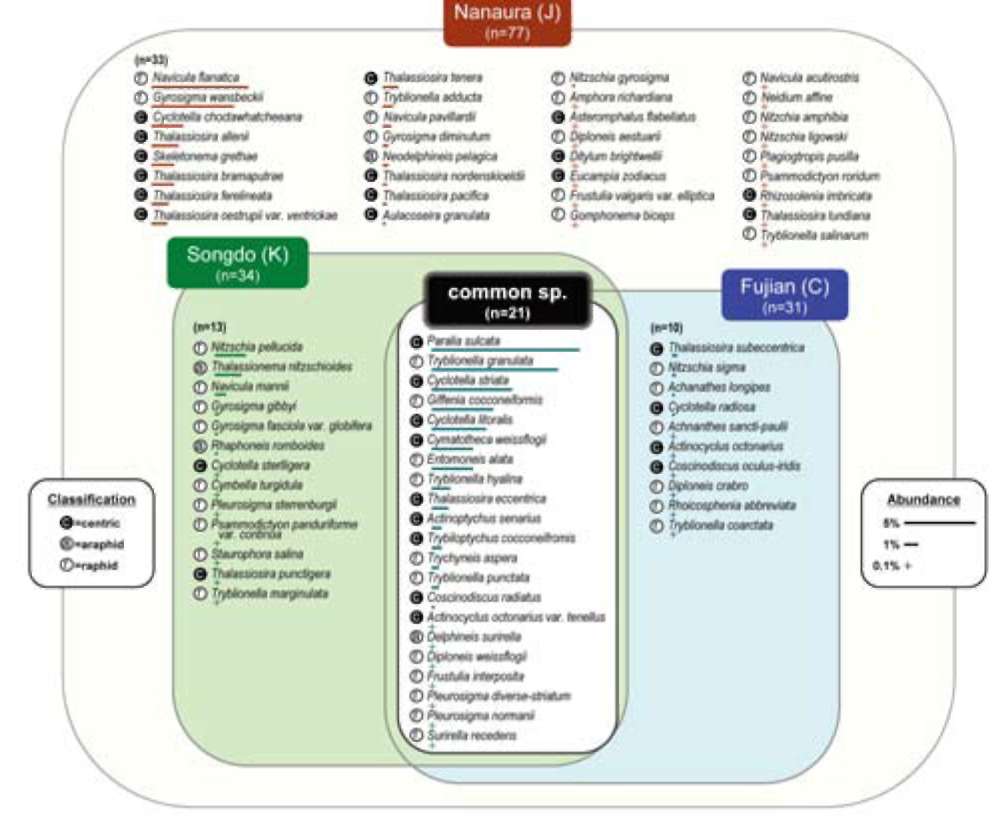
Dominant species in Northeast Asian tidal flats seem to be different from their European and North American counterparts, judging from previous reports as well as personal observations. For example, often missing in Korea and Japan are large motile species representing genera of e.g. Navicula, Nitzschia, Bacillaria, Cylindrotheca, Gy-rosigma, Pleurosigma, and Surirella (Hustedt, 1939, 1955, 1959; Brockmann, 1950; Hendey, 1964; Underwood, 1994; Witkowski et al., 2004), which often dominate European tidal flats. While the predominant species in Northeast Asia, accepting that data is limited, are reported as a large Haslea species (H. nipkowii) from a Japanese mudflat (Othsuka, 2005) and Fogedia and Navicula (personal data) from the Korean sand flat. To better understand their floral characteristics in terms of biogeography, we first tried to extract the typical and common species in Japan (Nanaura), Korea (Songdo), and China (Fujian), then address their global distribution.
|
||
|
In brief, the present study aimed to: 1) report the diatom flora of the Nanaura mudflat, Ariake Sea, along with each species' relative abundance and dimensional and micro-structural information based on light microscopic (LM) and scanning electron microscopic (SEM) observations; 2) examine and compare concurrently occurring species between Nanaura mudflat and Northeast Asia and their biogeographic distribution; and 3) extract and address the global distribution of typical and/or common Northeast Asian species.
|
||
|
MATERIALS AND METHODS
|
||
|
Sampling and observation
|
||
|
The top 0-0.5 cm of surface sediment was collected
|
||
|
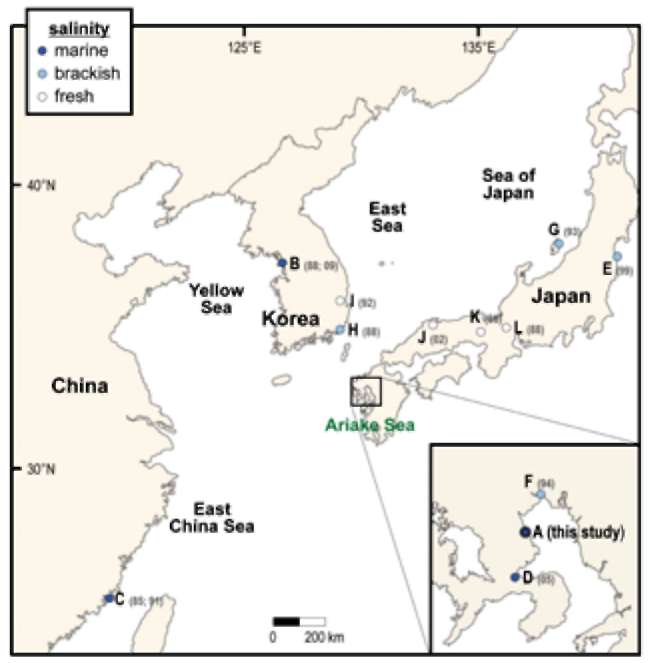
Figure 1. Map showing the study area and 11 other benthic habitats in Northeast Asia from which diatom flora literature were reviewed. A-D (
|
||
|
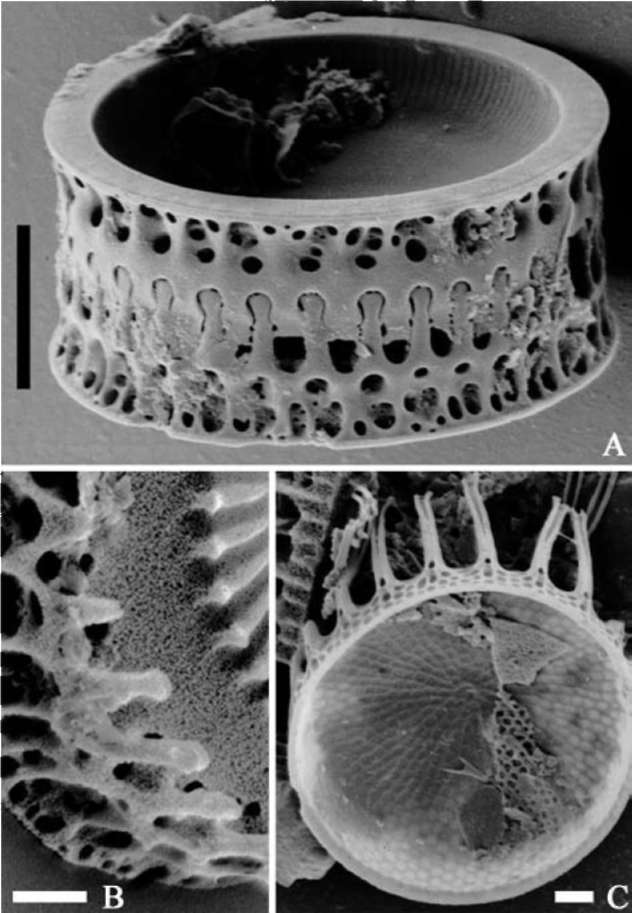
from the uppermost tidal flat during ebb tide at Nanaura (E 130。 10', N 33° 04'), Ariake Sea, Japan, on Oct. 2002 (Figure 1). Permanent slide cleaning and preparation involved a concentrated HCl and H2O2 treatment to remove calcium carbonate particles in the sediment and organic materials in the cells. Permanent slides were made using pleurax mountant. The specimens were then photographed under the Olympus BX60 LM with x60 Plan Apo objective lens, Zeiss Axioscop LM with x100 Achromat objective lens, and JEOL 6301F SEM (Figure 4). Under the LM observation (Figures 5-8), ca. 1,000 valves were counted and the relative abundance of each taxon was calculated.
|
||
|
Literature survey
|
||
|
A literature survey followed by comparative analysis were performed to gain a better understanding of the floral characteristics of Nanaura diatoms. Of major interest herein was to delineate the regional and habitual distribution of Nanaura diatoms across the various benthic habitats in Northeast Asia.
|
||
|
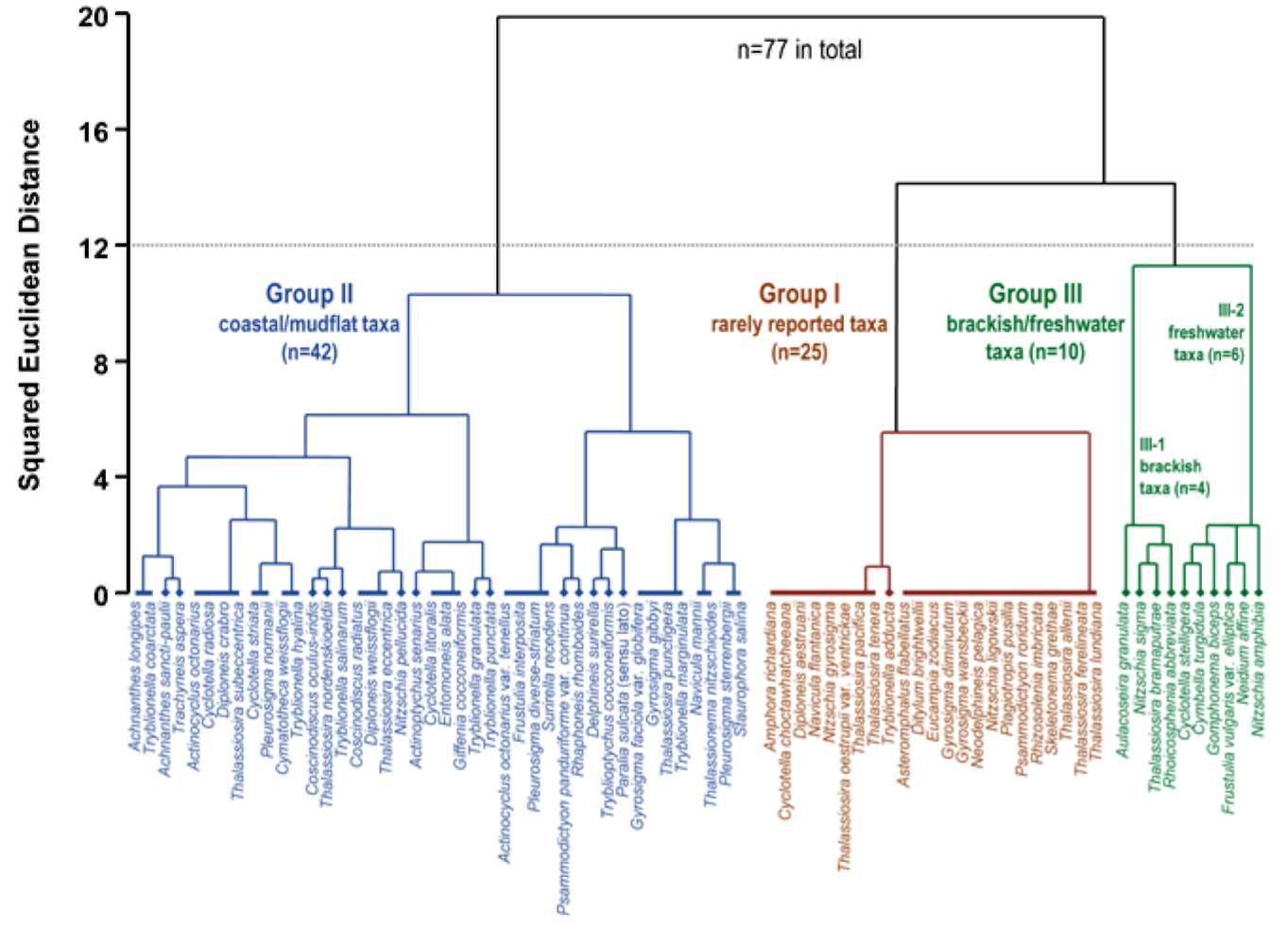
Diatom taxa identified to the species level only (n=77) were considered for the analysis. First, we collected all
|
||