90
Botanical Studies, Vol. 51, 2010
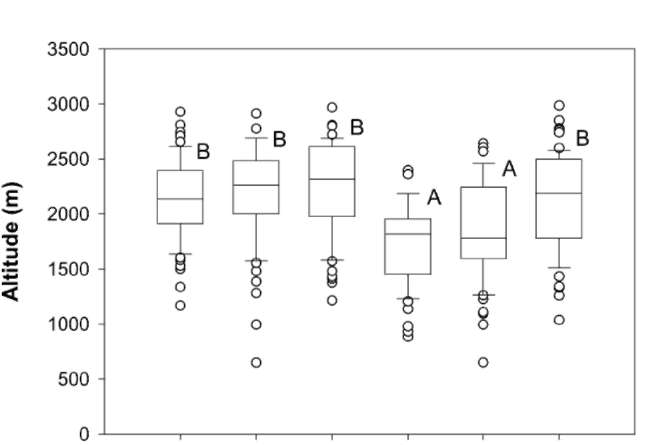
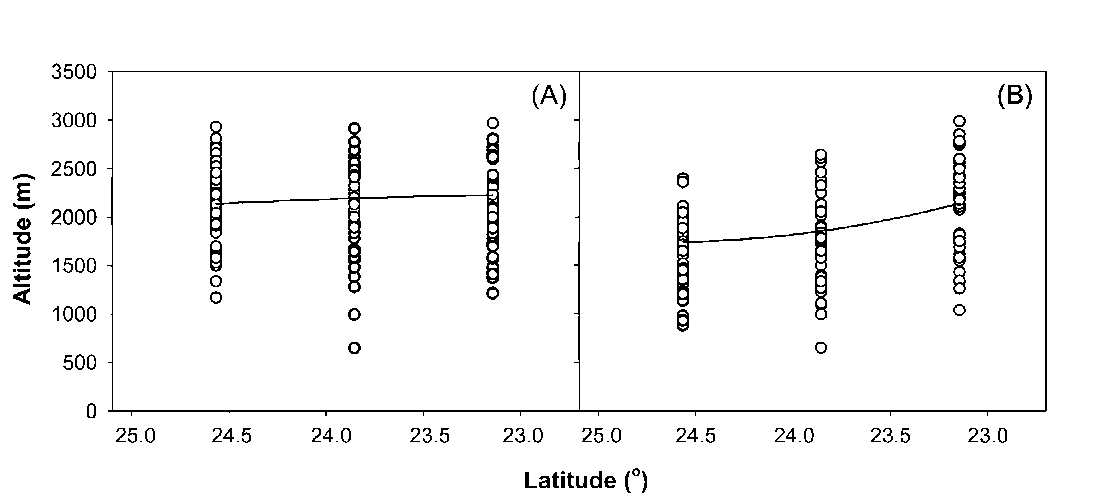
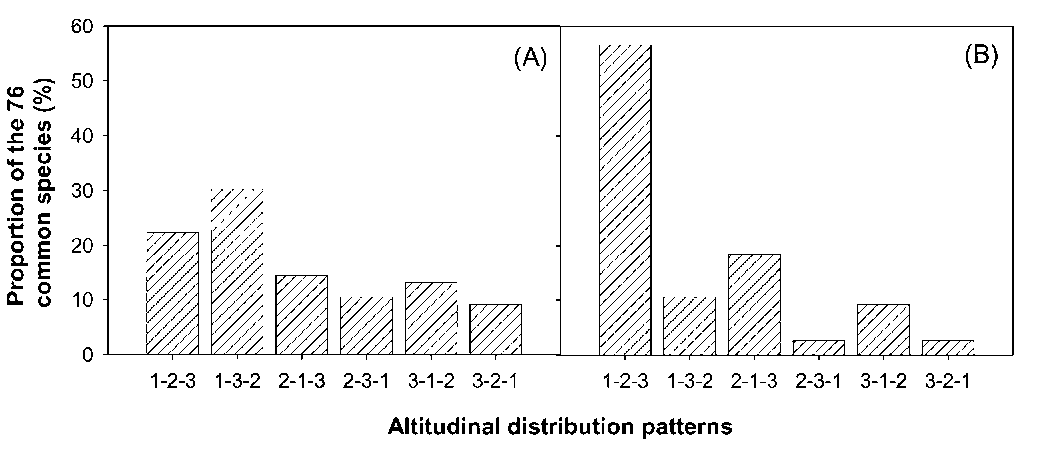
the marked descent of species and vegetation distribution in altitude has been documented (e.g. Hsieh et al., 1996; Lin et al., 2007). In the CMR main sections, where the ridges are higher than 2,000 m a.s.l. (Figure 1A), some ecologists have inferred that identical plant species or similar vegetations distribute highest in the central section and descend towards the both ends of the CMR (e.g. Su, 1984b). However, this is still open to question because the inference is made based on species distribution data collected from a small number of mountains on the west-
facing aspect of the CMR (e.g. Su, 1984b). The pattern
of species altitudinal distribution in Taiwan has to be re-examined based on data with relatively widely-distributed sampling plots.
Two mechanisms have been proposed to explain the altitudinal distribution patterns in Taiwan, including the heat retention mechanism of Massenerhebung (e.g. Su, 1984a, 1984b) and effects of monsoon (e.g. Hsieh et al., 1996). According to the heat retention mechanism, at a given altitude, temperature is lower on small mountains than on large mountains because smaller mountain mass tends to keep less heat from solar radiation (e.g. Holtmeier, 2003). Mountains are less massive and shorter at the north and south ends than in the main sections of the CMR (Figure 1). Consequently, the descent of vegetation altitudinal distribution at the two ends has been attributed to this mechanism because of relatively small mountain mass there (e.g. Su, 1984a, 1984b). It has been documented that effects of the winter northeast monsoon on species
distribution are pronounced (Sun et al., 1998). The winter northeast monsoon can markedly reduce local temperature by means of bringing in cold air, increasing cloudiness and wind chilling. Due to the topographic effects of the CMR, the winter northeast monsoon exerts more influences on local climate at the two ends than in the main sections of the CMR (Gallus, 2000). As a result, some ecologists attributed the descent of species distribution at the ends of the CMR to the effects of the winter northeast monsoon (e.g. Hsieh et al., 1996). Nevertheless, these hypotheses remain unexamined due to the lack of proper ground-based species distribution data.
The aims of the present study are to: 1) identify the altitudinal distribution patterns for plant species in the main sections of the CMR; and 2) recognize the most influential environmental factor in species altitudinal distribution.
MATERIALS AND METHODS
Study area
Taiwan Island (21。55'-25。20, N, 119。30'-122。00, E) is located at the western edge of the Pacific Ocean, 150 km to the southeast of the Mainland China and 300 km north of the Philippines. Total area of the island with length of 394 km and width of 140 km is 35,800 km2. The altitude ranges from 0 to 3,952 m. The CMR stretching from the north to the south tips of Taiwan (Figure 1) has over 300 summits higher than 3,000 m altitude.
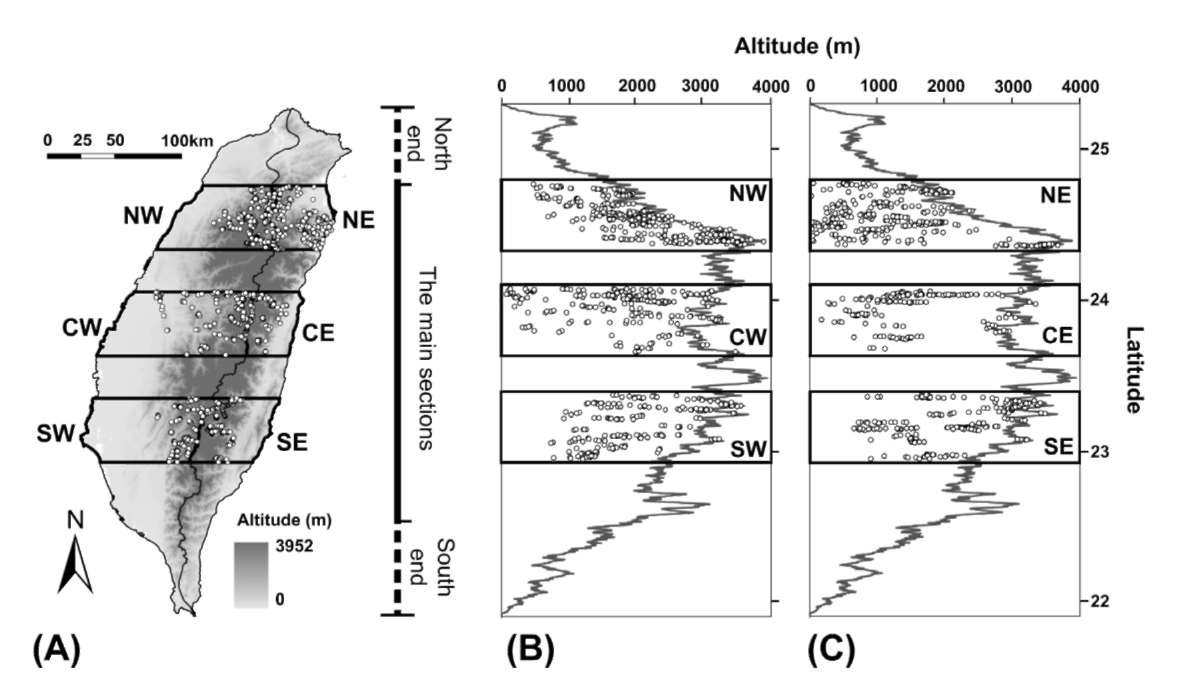
Figure 1. Map of Taiwan, showing the six sampling regions and the distribution of the sampling plots (open dots). (A) the horizontal distribution of the sampling plots; the altitudinal and latitudinal distribution of the sampling plots in the sampling regions (B) on the west-facing aspect and (C) on the east-facing aspect of the Central Mountain Range (CMR). The CMR stretches from the north to the south of Taiwan Island. The winding black solid line in Panel (A) indicates the main ridges of the CMR. The zigzag lines in Panels (B) and (C) indicates the altitude of the main ridges of the CMR along latitude. We only sample regions with at least one summit higher 3,000 m.