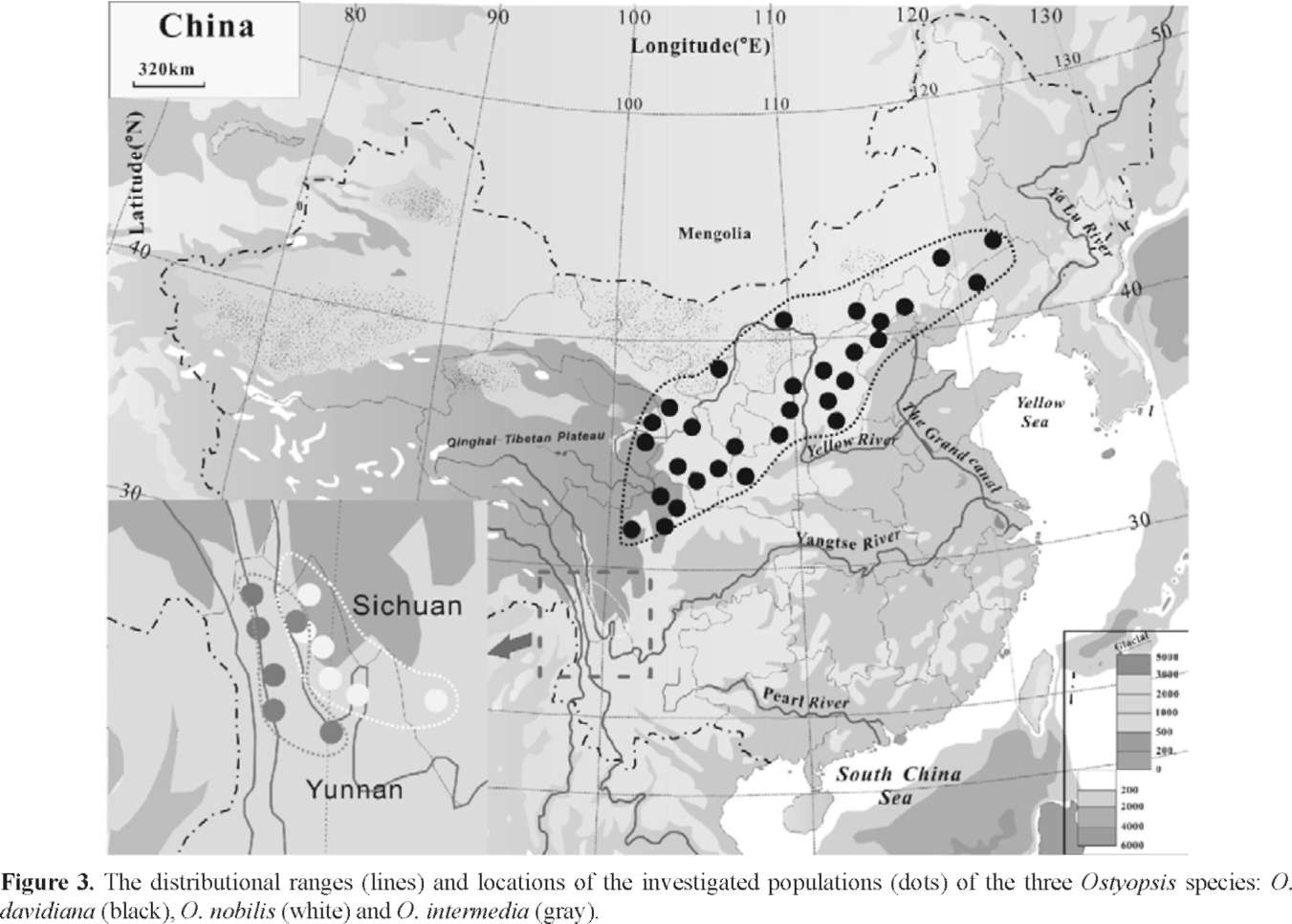
258
Botanical Studies, Vol. 51, 2010
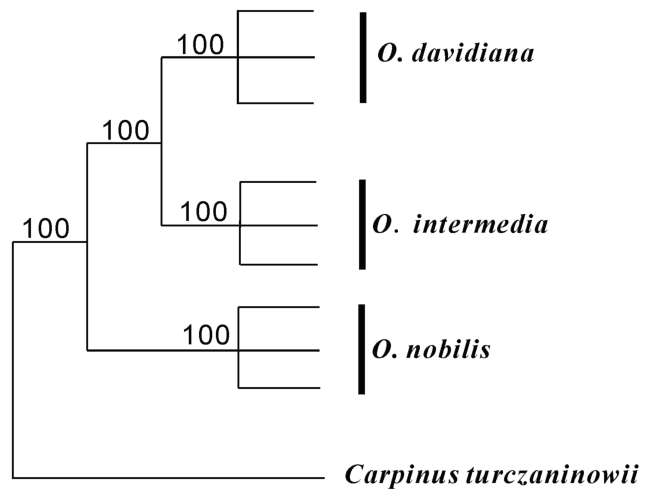
method of Doyle and Doyle (1987). The ITS regions of nrDNA were amplified with ITS primers 1 and 4 (White et al., 1990), then the PCR products were purified using a TIANquick Midi Purification Kit following the supplier's recommended protocol (TIANGEN). Sequencing reactions were performed with the PCR primers to cover the whole PCR segment using an ABI Prism BigdyeTM Terminator Cycle Sequencing Ready Reaction Kit. The reaction products were analyzed using an Applied Biosys-tems model 3130xl automated sequencer (Perkin Elmer Applied Biosystems). All sequences were submitted to GenBank (accession numbers: GQ250099 to GQ250101), then aligned using ClustalX version 1.81 (Thompson et al., 1997). Carpinus turczaninowii (AF081518) was selected
as an outgroup to root all sequenced individuals. The phy-logenetic relationships of the three species were assessed by maximum likelihood (ML), Neighbor-Joining (NJ) and maximum parsimony (MP) analyses using PAUP* 4.0b10. All gaps (indels) were coded as binary states (0 or 1). ML exhaustive search parameters were: simple addition of sequences of taxa with TBR branch swapping, MULTREES and COLLAPSE options on. MP analyses (equally weighted characters and nucleotide transformations) involved a heuristic search strategy with 100 replicates of random addition of sequences, in combination with ACCTRAN character optimization, MULPARS+TBR branch swapping and STEEPEST DESCENT options on. In these analyses we used bootstrap values (BS) to assess branch support.
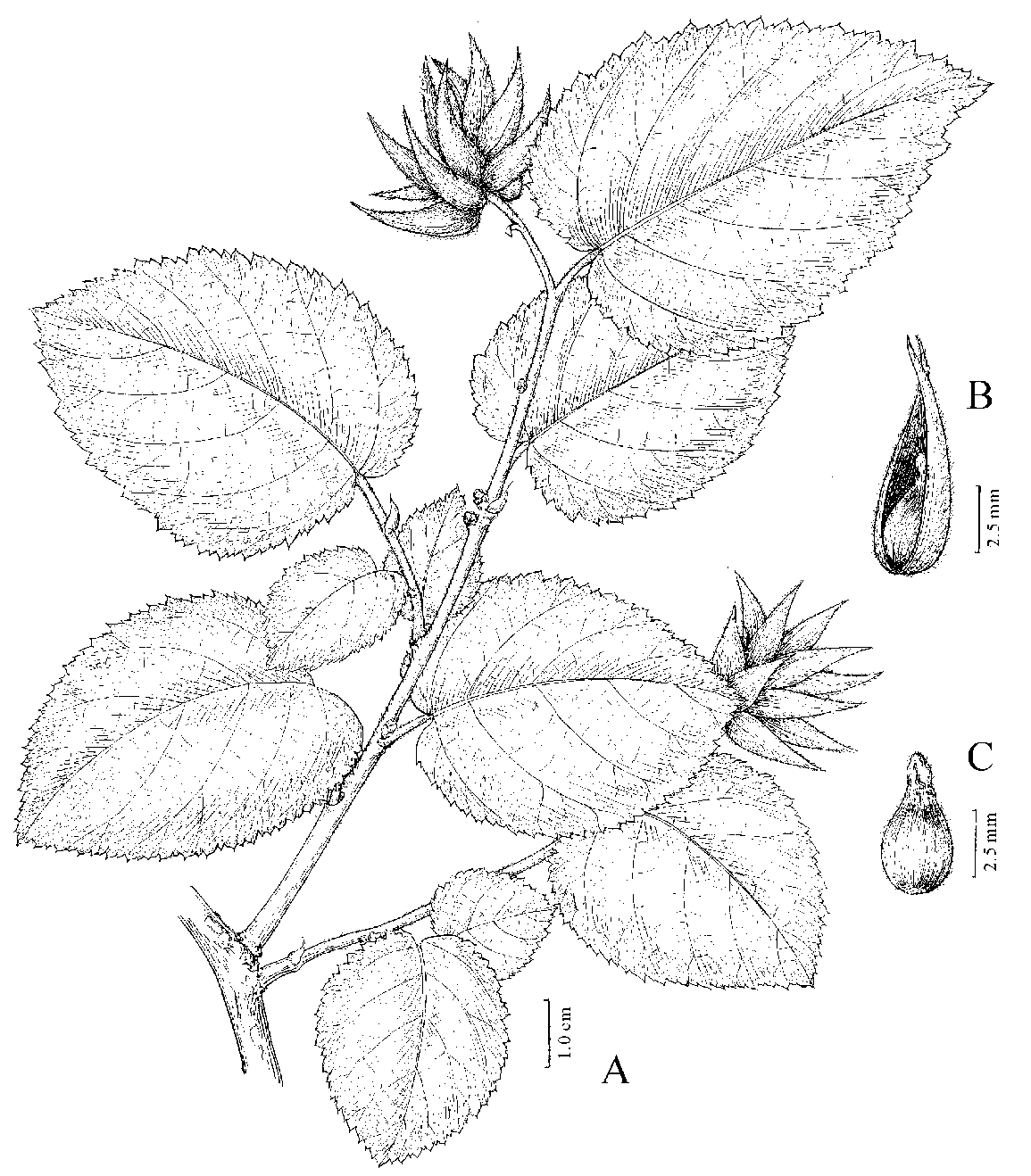
Figure 1. Ostryopsis intermedia B. Tian & J. Q. Liu A, fruiting branch; B, fruit; C, seed. Drawn by A. L. Li from B. Tian & J. Q. Liu 2007003 (LZU).