|
|
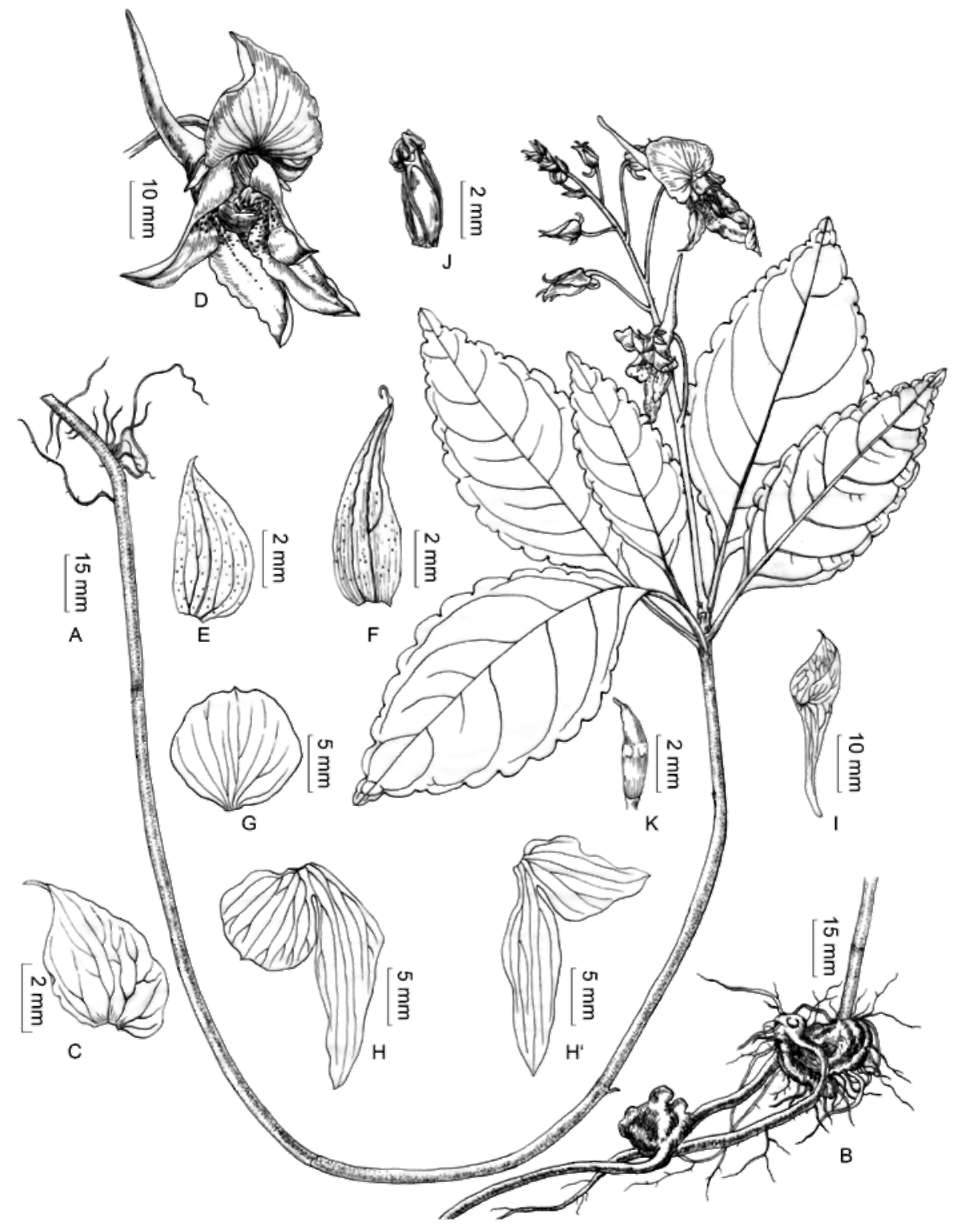
pink or pale pink; lateral united petals not clawed, left and right pairs unequal in size with the left pair often larger than the right pair, thus the flower being oblique and asymmetrical, pink or light pink, yellow with dense red dots at throat; upper petals of the lateral united petals of each pair unequal in size, orbicular to ovate, 8-12 x 10-16 mm, apex round or obtuse; lower petals of the lateral united petals oblong, oblique, incurved, 20-30 x 5-10 mm, much longer than the upper petals, distally ligulate. Filaments linear, swollen at upper part; anthers ovoid, apex obtuse. Ovary ovoid-oblong, glabrous. Capsule narrow-fusiform or clav-ate, 1-3 cm long, 5-8 mm wide; seeds 3-6, ca. 1 mm long, brownish.
Phenology. Flowering in July to September; fruiting in September to October.
Ecology. Under stories of sparse and dense broad-leafed forests on slopes, forest margins in moist places in valleys;
700-1,400 m.
Distribution. Sichuan, Southwestern China. It is so far known only from the type location and its vicinity, sparsely found in both front and rear hills of Qingchengshan Park.
Horticulture potential. This new species has large and showy flowers and was easy to grow in a garden due to its unique rhizome structure. Thus, it has great potential for horticultural applications.
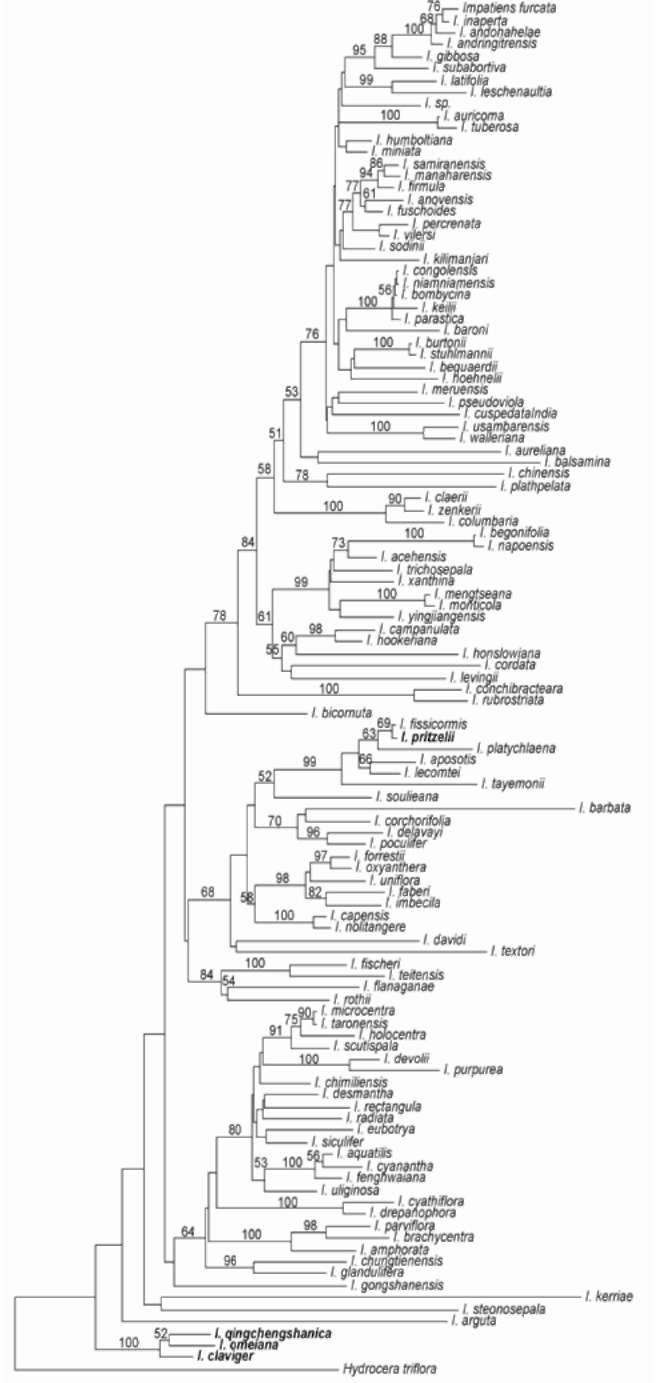
Phylogenetic Position of the New Species. In her doctoral dissertation, Song (2006) studied the molecular phylogeny of the family Balsaminaceae based on multiple DNA markers and revealed that this new species (represented as an unidentified specimen Isp2004-16), together with I. clavigera and I. omeiana, was nested in the most basal clade of the gene tree of the genus Impatiens. In order to further confirm the phylogenetic relationships of the new species, we revisited the phylogeny of the genus Impatiens. The DNA sequences of the internal transcribed spacers (ITS) of the nuclear ribosome DNA of the new species I. qingchengshanica (DNA obtained from the type specimen, GenBank accession HQ718764), its putative close relatives based on gross morphology, I. clavigera (voucher specimen AF23 deposited in NEU, sample collected by A. Favre from Fannipan, Sapa, northern Vietnam at the altitude 1,500 m on 15 September 2003; GenBank accession HQ718766), and I. omeiana (voucher specimen Yuan Y.-M. 2004-21 deposited in NEU, collected by Y.-M. Yuan from Emeishan Mountain, Sichuan, China at the altitude 1,200 m on 13 September 2004; GenBank accession HQ718765) were acquired and analyzed together with the previously published data matrix of the ITS sequences (Yuan et al., 2004). I. pritzelii, another putative close relative with similar rootstock, was already included in the previous study. The three newly-acquired ITS sequences were aligned with the original ITS matrix containing 112 taxa of Balsaminaceae (Yuan et al., 2004) using Clustal X (Thompson et al., 1997). A molecular phylogenetic tree was reconstructed by PAUP* v4.0b10 (Swofford, 2000) using the neighbor-joining method. Hydrocera triflora was used as the outgroup. Bootstrap clade support values
|
were calculated based on 1000 replicates of data applying the same neighbor-joining method. The resulted neighbor-joining tree, as shown in Figure 4, confirmed the previous results. The new species I. qingchengshanica was clustered together with I. clavigera and I. omeiana as a highly supported clade (bootstrap 100%) that was placed as the most basal clade in the ITS molecular tree of the genus Impatiens. Impatiens pritzelii, a species having rootstocks very similar to our new species, was nested inside the ITS tree and distantly related to our new species (Figure 4). However, the resolution of the relationships among the main clades did not obtain strong bootstrap support for ITS sequences. This could be due to the relative high level of homoplasy in ITS sequences because other published data of chloroplast DNA sequences obtained much higher clade supports for the same relationships (Song, 2006).
Many species of Impatiens, especially perennial ones, have well-developed rhizomes (Grey-Wilson, 1980); however, modified rootstock is uncommon in Balsami-naceae. Only a few species are described having tubers or nodular rhizomes, these are: I. tuberosa H. Perrier from Madagascar, I. cinnabarina Grey-Wilson from Tanzania, I. omeiana from Sichuan, China, and I. pritzelii from Hubei and Chongqing, China. These species are geographically isolated from each other. In the molecular phylogenetic tree, they are also distantly related, suggesting that their rootstock, adapted for storage, may have undergone multiple independent origins and parallel evolution within the genus. The fleshy and enlarged tuber or nodular knots of Impatiens species are probably adaptations to the seasonal dry conditions of their habitats.
Acknowledgements. The field trips were financially supported by the Swiss National Science Foundation 3100A0-102165 granted to Y.-M. Yuan, and the Doctoral Program of Higher Education of China (20060558090) granted to X.-J. Ge. Ms. Yun-Xiao Liu kindly prepared the illustra-tion for us. We are much indebted to Dr. Jason Grant, Dr. Qin-Er Yang and Dr. Yun-Fei Deng who critically read the manuscript and kindly helped us in preparing the Latin di-agnosis.
LITERATURE CITED
Chen, Y.L. 2001. Flora Reipublicae Popularis Sinicae. Tomus, Vol. 47(2). Science Press, Beijing.
Cong, Y.Y., K.M. Liu, X.Z. Cai, and S.Z. Tian. 2008. Impatiens fugongensis (Balsaminaceae), a new species from Yunnan, China. Bot. Stud. 49: 161-165.
Grey-Wilson, C. 1980. Impatiens of Africa. A. A. Balkema, Rotterdam, Netherlands.
Huang, S.-H., Y.-M. Shui, and W.H. Chen. 2003. New taxa of
Impatiens from Yunnan. Acta Bot. Yunnan. 25: 261-280.
Janssens, S., K. Geuten, Y.-M. Yuan, Y. Song, P. Kupfer, and E. Smets. 2006. Phylogenetics of Impatiens and Hydrocera (Balsaminaceae) Using Chloroplast atpB-rbcL Spacer Se-
|
|